드디어 DNA의 조직화를 다룰 차례다.
이 포스팅을 읽는 독자라면 아마도 염색체(Chromosome)라는 용어와 염색체의 X 모양 구조가 익숙할지도 모른다. 염색체는 DNA가 고도로 응축한 결과물이다. 그렇게 얇고 긴 DNA가 어떻게 눈에 보이는 염색체로 응축될까?
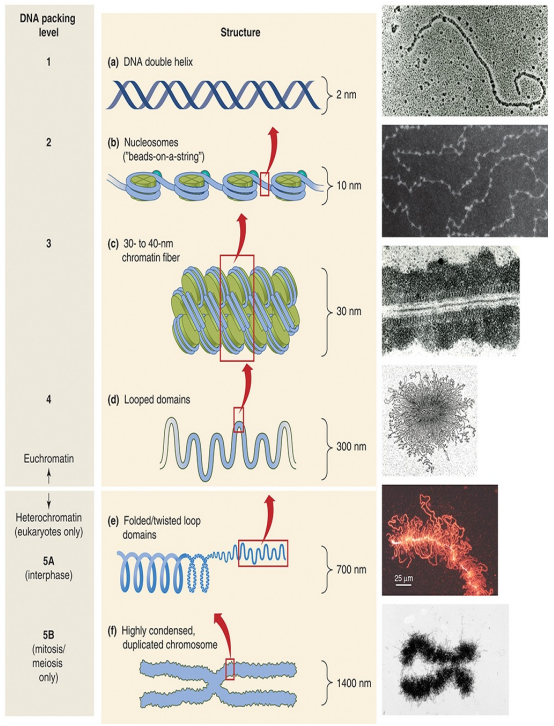
DNA의 응축 과정을 DNA의 조직화(Organization)라 부르며, 세포(특히 진핵세포)는 총 다섯 단계의 조직화를 이용한다. 단계를 하나씩 알아보자.
1. 첫 번째 조직화 : 이중 나선(Double helix)의 형성
앞선 글에서 언급했듯, DNA는 단일 가닥으로 존재하지 않는다. 단일 가닥은 세포 내에서 안정성이 떨어지기 때문이다. 대신 매우 안정적인 이중 나선(Double helix) 구조를 형성한다.

DNA 두 가닥을 연결하는 힘은 염기끼리의 수소 결합(Hydrogen bond)이다. 이때 염기는 항상 상보적(Complementary)으로 쌍을 이룬다. 아데닌은 항상 티민과 수소 결합을 이루고, 사이토신은 구아닌과 수소 결합을 이룬다. (이하 A, T, G, C로 통칭)
또한 A와 T는 2개의 수소 결합, G와 C는 3개의 수소결합을 이룬다. 때문에 G-C 결합이 A-T 결합보다 조금 더 강하다.
상보적으로 결합한 두 가닥의 DNA는 서로 방향이 반대인 역평행(Antiparallel) 관계이다. 즉, 한쪽 가닥이 3'에서 5' 방향이라면, 다른 가닥은 5'에서 3'방향이다.

연결된 두 가닥의 DNA는 감기면서 나선 구조를 형성한다. 이때 DNA 나선의 방향은 오른 나선(right-handed) 방향으로, 엄지 손가락을 나선의 진행 방향으로 향했을 때 나머지 손가락이 굽혀지는 방향으로 감긴다.

DNA의 이중 나선 구조는 틈의 길이가 일정하지 않다. 즉, 항상 넓은 틈과 짧은 틈이 교대로 존재하는데, 이때 각각을 넓은 홈(Major groove)과 좁은 홈(Minor groove)이라 부른다.
넓은 홈과 좁은 홈이 반복되는 주기, 즉 나선이 한 바퀴 도는 주기는 3.4 nm(또는 34 Å)이다. 또한 DNA의 폭은 약 2 nm(또는 20 Å)이다.

DNA 이중 나선 구조는 DNA가 존재하는 환경에 따라 다양한 형태(Form)를 가질 수 있다. 우리가 가장 일반적으로 접하는 DNA는 B 형태(B from)이다. 세포 내부와 같은 상대적으로 높은 습도 환경에서 DNA는 B 형태를 선택한다.
반면, 실험 환경과 같은 상대적으로 낮은 습도 환경에서 DNA는 A 형태(A form)를 선택한다.
A, B 형태 모두 앞서 언급했던 오른 나선 방향으로 감겨있는데, 세 번째 형태인 Z 형태(Z form)는 특이하게도 왼 나선(Left-handed) 방향으로 감겨 있다. Z 형태의 역할은 명확히 밝혀지지는 않았으나, 아마도 전사 과정에서 필요한 DNA 구역이 풀릴 때, DNA 전체의 과한 감김(Supercoiling)을 방지하기 위함이라 추측된다.
2. 두 번째 조직화 : 뉴클레오솜(Nucleosome)의 형성
세포의 입장에서, 이중 나선을 형성한 DNA(이하 DNA로 통칭)는 그대로 두기에는 너무 길다. 사람의 한 세포에 들어있는 DNA의 길이는 약 1.8 m에 달한다고 하니 말이다.
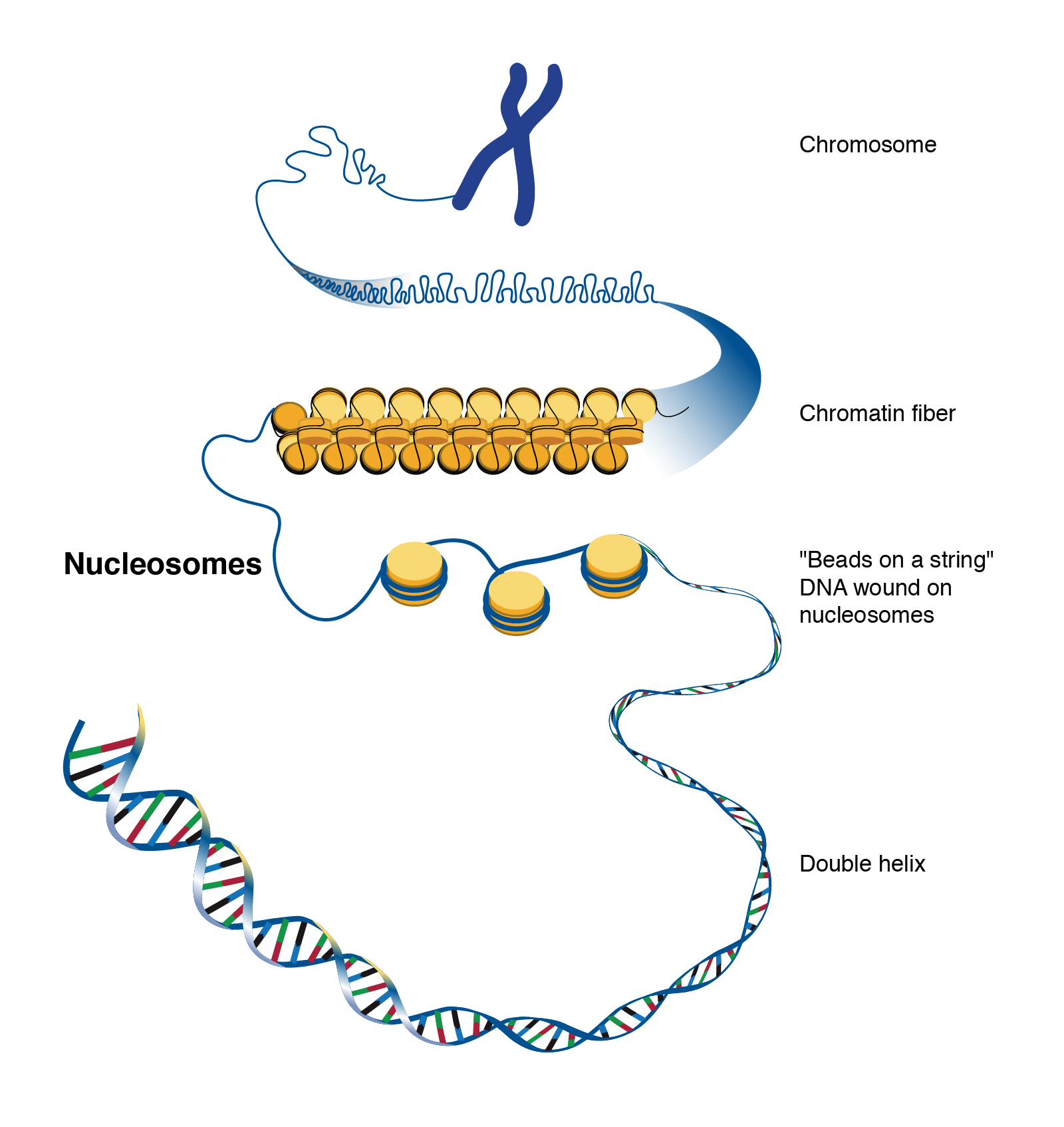
긴 끈을 감아 정리하듯, DNA도(특히 진핵세포에서) 특정 구조물에 감겨 조직화되는데, 이 특정 구조물을 히스톤(Histone), 혹은 히스톤 단백질(Histone protein)이라 부른다. 히스톤은 전체 염색체의 질량 중 50%를 차지할 정도로 DNA의 조직화에 있어 중요한 역할을 담당한다.
히스톤은 구형의 단백질 복합체다. DNA는 히스톤 주위로 약 1.7 바퀴(167 bp)만큼 감겨 있다. 히스톤과 감겨 있는 DNA를 한데 묶어 뉴클레오솜(Nucleosome)이라 부른다.
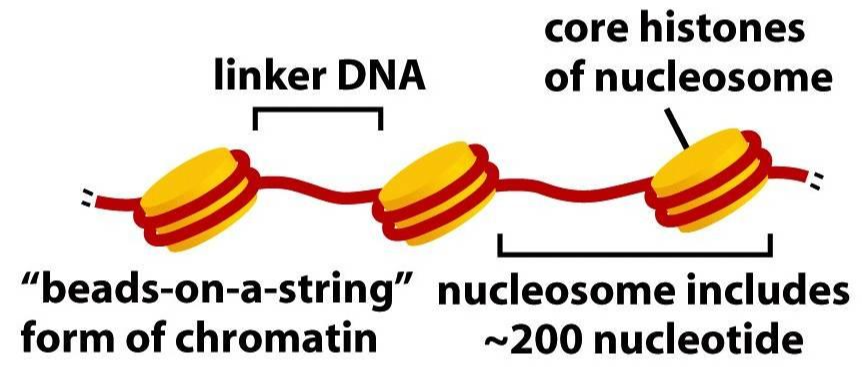
뉴클레오솜은 이웃한 뉴클레오솜과 일정 길이의 DNA를 두고 떨어져있다. 긴 줄에 구슬이 간격을 두고 이어진 그림을 상상해 보면 좋을 것이다. 두 뉴클레오솜 사이의 DNA를 연결자 DNA(Linker DNA)라고 부른다.
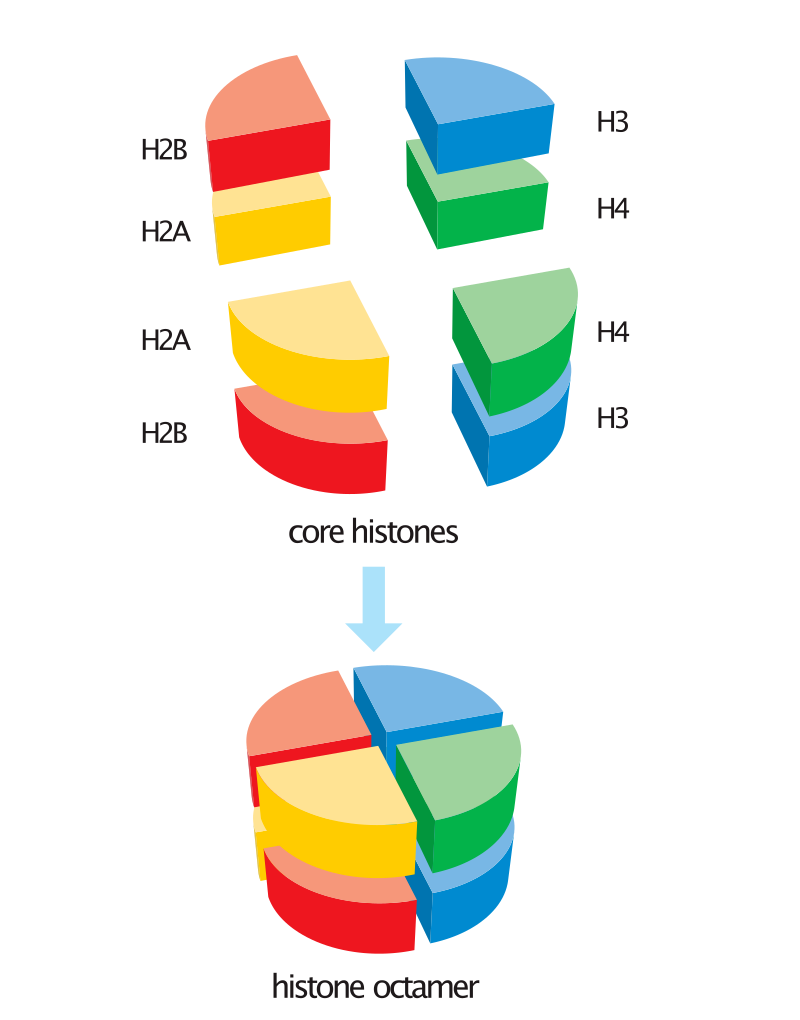
뉴클레오솜의 히스톤은 H2A, H2B, H3, H4 네 개의 히스톤 단백질이 모여 하나의 단위체를 만들고, 그러한 단위체가 두 개 모여 구성된다. 즉, 히스톤은 8량체(Octamer)이고, 이를 뉴클레오솜 코어 입자(Nucleosome core particle, NCP)라고 부른다.
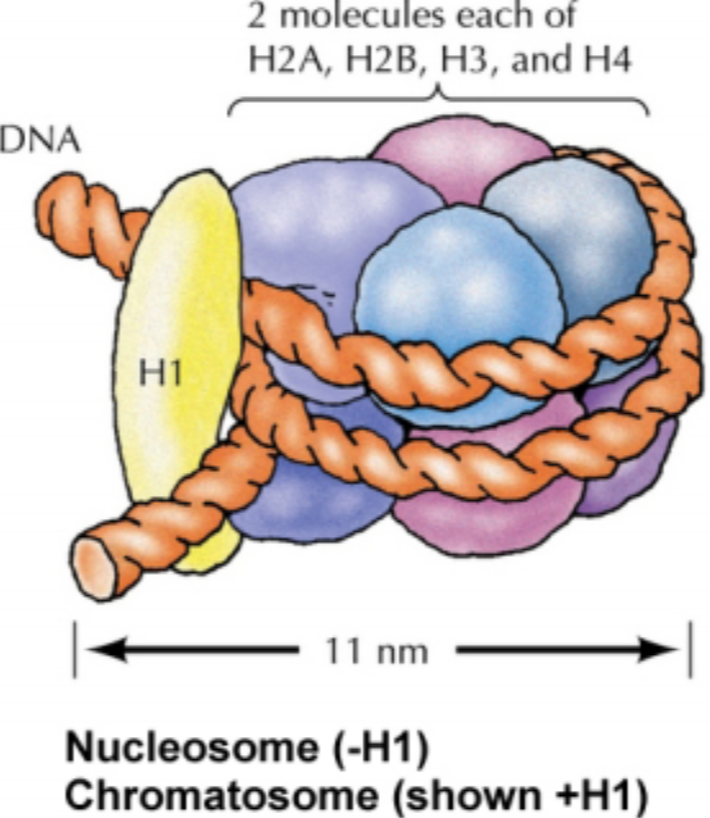
감긴 줄이 풀리지 않게 핀을 박아두듯, 히스톤에 감긴 DNA 또한 이용하지 않을 땐 풀리지 않게 고정되어야 한다. 이때 연결자 단백질(Linker protein)인 H1이 사용된다. 이름에서 알 수 있듯 NCP를 구성하는 단백질과 같은 족에 속한다. 뉴클레오솜에 H1이 결합하여 본질적으로 불활성화(Deactivating)된 구조를 크로마토솜(Chromatosome)이라 부른다.
크로마토솜이 되어 불활성화된 DNA를 이용하고 싶을 때, 세포는 SWI/SNF 단백질 족(Family)을 이용한다. 이들은 ATP를 사용하여 크로마토솜에서 연결자 단백질인 H1을 제거해 이용 가능한 뉴클레오솜으로 되돌린다.
3. 세 번째 조직화 : 30-nm 섬유(30-nm fiber)의 형성
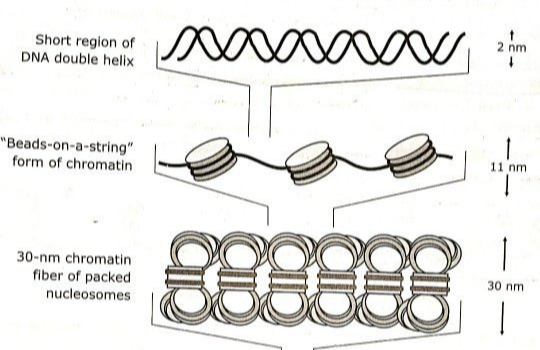
길게 연결된 여러 개의 뉴클레오솜은 한번 더 뭉쳐 30-nm 섬유(30-nm fiber)―원핵생물에서는 40-nm 섬유―를 형성한다.
뉴클레오솜이 뭉치는 원인은 히스톤 단백질의 전하 때문이다. 뉴클레오솜 코어 입자의 H4는 음전하를 갖는데, 이것이 H2A/H2B의 양전하와 상호작용하며 뉴클레오솜을 응축되게 만든다. 따라서 세 번째 조직화, 30-nm 섬유의 형성은 자발적으로 일어나는 반응이다.
세 번째 조직화의 결과로 이중 나선 DNA는 약 42배 만큼 응축된다.
4. 네 번째 조직화 : 루프 도메인(Loop domain)의 형성
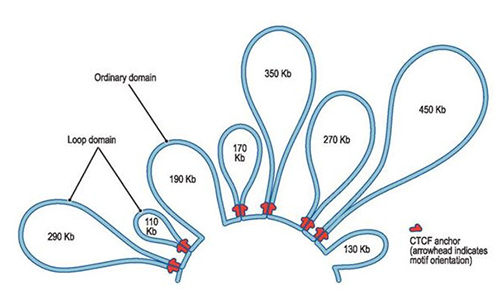
30-nm 섬유는 다시 특정 길이만큼 감겨서 '포장'되는데, 이렇게 포장된 영역을 루프 도메인(Loop domain)이라 부른다.
루프 도메인의 형성에는 콘덴신(Condensin), 코헤신(Cohesin)과 같은 다양한 단백질이 관여하는데, 이에 대해서는 세포생물학이 다루는 범위를 벗어나므로 자세히 다루지 않고, 차후 분자생물학 포스팅에서 이야기해 보도록 하자.
원핵생물의 염색체는 4단계 조직화까지만 가능하다. 그리고 DNA 이중 나선 구조에서부터 루프 도메인에 이르기까지 네 단계 조직화가 진행된 염색질을 진정염색질(Euchoromatin)이라 부른다.
5. 다섯 번째 조직화 : 이질염색질(Heterochromatin)의 형성
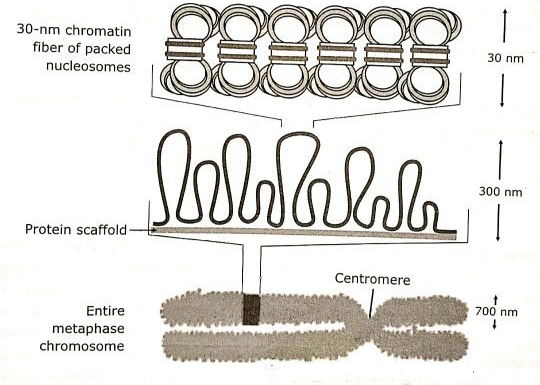
루프 도메인이 조금 더 복잡하게 꼬이면 본질적으로 이용이 불가능한 이질염색질(Heterochromatin)이 형성된다. 이질염색질은 오직 진핵생물의 핵에서만 발견된다.
이질염색질은 응축 정도에 따라 다시 두 단계로 나눌 수 있다.
세포 주기의 간기 동안, 이질염색질은 풀린 상태로 존재한다. 그러다가 세포 분열이 시작되면 이질염색질은 염색체(Chromosome)로 한 번 더 응축된다. 즉, 우리가 흔히 접했던 염색체는 아주 많이 응축된 상태의 DNA라고 볼 수 있다.
6. DNA의 변형(Modification of DNA)
조직화 이외에도, 세포는 다양한 방법으로 DNA를 변형(Modify)해가며 이용한다.
DNA 변형(DNA modification)은 분자생물학의 주된 토픽 중 하나인데, 세포생물학 포스팅에서는 DNA의 조직화와 관련된 약간의 내용만 간단하게 다뤄보도록 하자.
1) DNA의 메틸화
대표적인 DNA 변형 중 하나는 DNA 메틸화(DNA methylation)다. 메틸화는 특정 부분의 유전자를 침묵(Silencing)시켜 발현되지 못하도록 만든다.
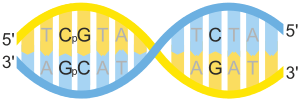
DNA 메틸화는 원핵세포에서는 아데닌, 진핵세포에서는 사이토신과 구아닌에서 일어난다. 특히 진핵세포에서 DNA 메틸화가 일어나는 곳은 구아닌과 사이토신이 인접한 채로 반복되는데, 이 서열의 사이토신을 약자로 CpG라고 부른다. 약자에서 p는 두 염기가 인산다이에스터결합을 이루고 있다는 것을 뜻한다.
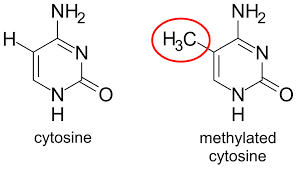
특정 유전자의 CpG가 메틸화되면, 원래 그곳에 결합해야 할 전사 인자(Transcription factor, TF)가 제대로 결합하지 못하고, 결과적으로 유전자의 발현이 억제된다.
2) 히스톤 단백질의 변형
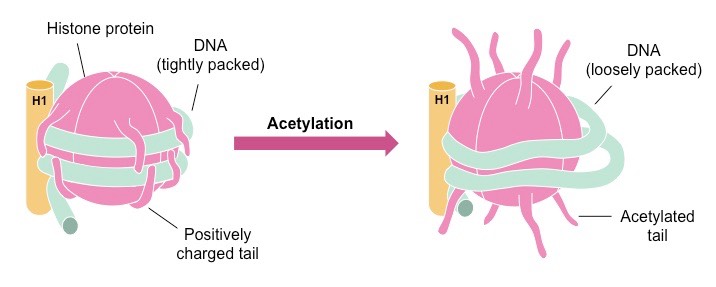
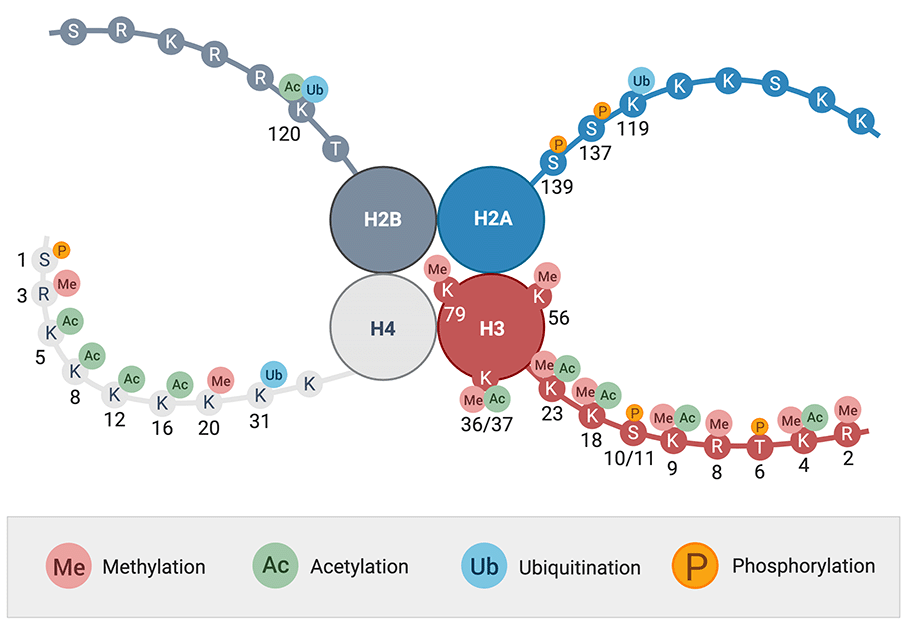
두 번째 DNA 변형은 DNA의 두 번째 조직화에 작용한다. 히스톤 단백질은 꼬리(Tail)를 가지고 있어, 다양한 조절 인자들이 결합할 수 있다. 조절 인자에는 대표적으로 아세틸기(Acetyl group, Ac), 메틸기(Methyl group, Me), 인산기(Phosphate group, P)가 있다.
드디어 DNA 파트의 마지막 글이다.
다음 주제는 단백질이다. 단백질이 무엇이고 세포 내에서 어떤 기능을 갖는지 공부해보도록 하자.
'세포생물학' 카테고리의 다른 글
| [세포생물학] DNA의 구조 (Structure of DNA) (0) | 2024.04.10 |
|---|---|
| [세포생물학] DNA와 RNA (DNA and RNA) (1) | 2024.04.06 |
| [세포생물학] 생명 (The Life) (2) | 2024.03.29 |
| [세포생물학] 세포생물학 (Cell Biology) (1) | 2024.03.29 |