
대부분의 조직에서 대사 에너지는 거의 전적으로 포도당에 의존한다. 그래서 외부로부터의 포도당 유입이 없을 때 포도당이 고갈될 경우 개체는 심각한 손상을 입을 수밖에 없다. 때문에, 비탄수화물 전구체(피루브산 등)로부터 포도당을 합성하는 포도당신생성(Gluconeogenesis) 경로가 모든 종에서 발달되어왔다.
1. 포도당신생성 개요
포도당신생성 반응은 당분해 과정처럼 모든 조직과 모든 종에서 본질적으로 동일하고, 반응의 전구체로 쓰이는 물질 정도만 조금씩 다르다. 동물, 그중에서도 특히 포유동물의 간세포에서 일어나는 포도당신생성은 당분해 반응의 역과정에 가장 가깝다. 이번 글에서는 가장 기본적인 포도당신생성 반응에 대해 알아보도록 하자. 두 과정을 함께 살펴보는 것이 공부하는 데 있어 도움이 많이 될 것이다.

2. 포도당신생성의 우회경로
당분해 과정이 그랬듯, 포도당신생성 과정도 10단계로 나누어진다. 그중 7개 단계는 당분해의 완전한 역반응이다. 하지만, 나머지 3개 단계는 매우 큰 음의 ΔG를 갖고 있어 비가역적이다. 때문에, 포도당신생성에서는 세 가지 우회 반응을 갖고 있다.
1) 첫 번째 우회 반응 : 피루브산 → 인산엔올피루브산
피루브산을 인산엔올피루브산(PEP)으로 인산화시키는 반응은 옥살아세트산(Oxaloacetate)이라는 중간체를 필요로 한다. 우선, 피루브산 카복실화효소(Pyruvate carboxylase)가 피루브산을 중탄산염(Bicarbonate)과 결합시켜 옥살아세트산으로 변환한다.
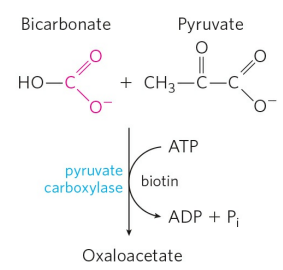
피루브산 카복실화효소는 바이오틴(Biotin)이라는 '중탄산염 운반체'를 필요로 한다. 피루브산 카복실화효소와 바이오틴의 기전에 대한 내용은 차후 시트르산 회로(TCA 회로)를 공부할 때 보다 자세히 다루도록 하고, 피루브산 카복실화효소는 한 분자의 ATP를 소모한다는 점만 기억하면 된다.

그 후, 인산엔올피루브산 카복시인산화효소(Phosphoenolpyruvate carboxylkinase)가 옥살아세트산을 인산엔올피루브산(PEP)으로 변환시킨다. 옥살아세트산은 4-탄소 화합물이고, 피루브산은 3-탄소 화합물임을 기억하라. 이 반응에서 방출되는 CO2는 앞서 피루브산을 옥살아세트산으로 바꿀 때 첨가한 중탄산염과 동일한 분자다.
옥살아세트산의 변환은 GTP를 소모한다. GTP는 에너지 수율을 계산할 때 ATP 한 분자와 동등하게 취급한다. 즉, 피루브산을 인산엔올피루브산으로 바꾸는 데 벌써 두 분자의 ATP를 사용한 셈이다. 이는 인산엔올피루브산이 높은 에너지를 가진 분자이기 때문이다.
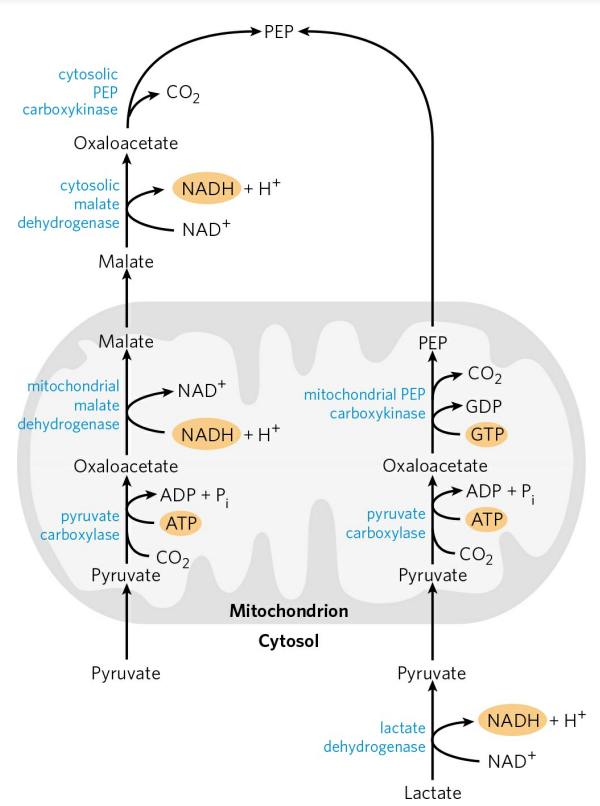
중간체로 옥살아세트산을 거치는 중요한 이유가 있다. 피루브산은 세포질보다 미토콘드리아에 풍부하다. 하지만 포도당신생성은 세포질에서 일어난다. 따라서, 피루브산을 포도당으로 재생시킬 때 피루브산을 세포질로 운반하는 과정이 필요하다. 하지만 미토콘드리아 막에는 피루브산을 수송하는 수송 단백질이 없다.
피루브산은 미토콘드리아 내에서 옥살아세트산으로 일차적으로 변환된 후, 미토콘드리아 말산 탈수소효소(Malate dehydrogenase)(세포질에 존재하는 동종 효소(세포질 말산 탈수소효소)와 구별하기 위함)에 의해 말산(Malate)으로 환원된다.
$ Oxaloacetate + NADH + H^+ \rightarrow Malate + NAD^+ $
이 반응은 ΔG가 0에 가까워 쉽게 양방향으로 반응이 일어난다. 말산은 미토콘드리아 내막 수송체를 통해 세포질로 운반되고, 세포질로 운반된 말산은 세포질에서 다시 옥살아세트산으로 산화된다.
피루브산을 옥살아세트산의 형태로 세포질로 수송하는 것 이외에도, 이 반응은 미토콘드리아의 NADH를 세포질로 운반하는 부가적인 기능(정확히는 운반이 아니라 세포질 NADH의 재생이다.)도 수행한다. 포도당신생성은 당분해의 역과정이므로 NADH를 필요로 한다. (당분해 과정은 NAD+를 필요로 한다.) 이때 필요한 NADH가 말산의 수송 과정에서 공급된다. 나중에 배우겠지만, 말산-옥살아세트산 변환은 미토콘드리아-세포질 간 [NADH]/[NAD+] 비율을 조정하는 유용한 기전 중 하나이다.
아직 다루지 않았지만, 혐기성 젖산 대사가 우세한 근육과 같은 저산소 환경에서는 세포질 내 NADH가 풍부해진다. 이때는 굳이 옥살아세트산이 말산으로 전환되어 세포질로 나갈 필요 없이, 미토콘드리아 내에서 바로 인산엔올피루브산으로 변환된다.
2) 두 번째 우회 반응 : 1,6-이중인산 과당 → 6-인산 과당
6-인산 과당의 인산화는 인산 과당 인산화효소-1(PFK-1)에 의해 이뤄진다. 반면, 1,6-이중인산 과당의 탈인산화는 과당 1,6-이중인산염분해효소-1(Fructose 1,6-bisphosphatase-1, FBPase-1)에 의해 이뤄진다. 이때 1번 탄소 인산의 가수분해가 일어난다.
3) 세 번째 우회 반응 : 6-인산 포도당 → 포도당
포도당의 인산화는 육탄당인산화효소에 의해 이뤄진다. 반면, 6-인산 포도당의 탈인산화는 포도당 6-인산염분해효소(Glucose 6-phosphatase)에 의해 이뤄진다. 이 효소는 간과 신장, 소장의 내피세포에만 존재해 당분해가 일어나는 다른 조직에서 6-인산 포도당의 가수분해가 일어나지 않도록 한다.
2), 3) 반응이 우회반응으로 진행되는 이유는 반응의 효소와 관련이 있다. 두 반응 모두 당분해에서는 ATP를 소모해 인산화시키는 '인산기 전달 효소(Kinase)'가 관여한다. 때문에, 이것의 역과정을 그대로 하기 위해서는 ADP를 ATP로 재생시켜야 한다. 하지만 1,6-이중인산 과당 및 6-인산 포도당은 ATP를 재생시킬 수 있을 정도로 고에너지 인산 화합물이 아니다. 따라서 포도당신생성의 두 탈인산화 반응은 우회 경로로 진행된다.
3. 포도당신생성의 결과
결과적으로, 포도당신생성의 반응식은 다음과 같다.
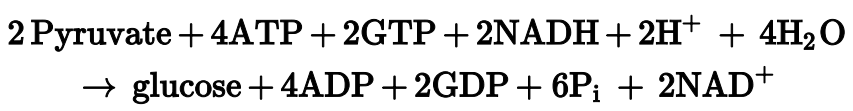
당분해 과정에서 두 분자의 ATP가 생성된다는 점을 떠올리자. 포도당신생성은 당분해의 역과정보다 네 분자의 ATP를 더 소모한다. 이는 포도당신생성의 비가역성을 확실하게 하기 위함이다. 당분해의 ΔG가 -63 KJ/mol 인 것을 고려하면, 포도당 신생성이 단순히 당분해의 역과정이라면 ΔG가 매우 큰 가역 반응이 된다. 실제로 위와 같은 우회 경로 때문에 포도당신생성의 ΔG는 -16 KJ/mol, 본질적으로 비가역적인 반응이 된다.
피루브산을 전구체로 하는 포도당신생성은 다양한 대체 경로가 가능하다. 대표적으로 첫 번째 우회경로의 중간체로 이용되는 옥살아세트산은 동시에 시트르산 회로의 중간체 중 하나이다. 따라서, 시트르산 회로의 모든 중간체는 이론적으로 포도당신생성에 이용될 수 있다. 또한, 대부분의 아미노산은 시트르산 회로의 중간체 혹은 피루브산으로 최종 대사되므로, 아미노산도 포도당신생성의 전구체로 이용될 수 있다. 이처럼 포도당신생성 경로로 대사 가능한 물질들을 통틀어 포도당생성(Gluconic) 물질이라 부른다.
단, 탄소 화합물과 아미노산과는 달리, 동물 세포에서 지방산은 포도당생성 물질이 될 수 없다. 나중에 배우겠지만, 지방산의 이화로 생기는 아세틸-CoA는 포도당으로 대사되는 경로가 존재하지 않는다. 자세한 내용은 나중에 지방산의 이화에 대해 다룰 때 보다 자세히 이야기하도록 하자.

'생화학 > 생화학 (하)' 카테고리의 다른 글
| [생화학] 발효 (Fermentation) (0) | 2022.07.31 |
|---|---|
| [생화학] 인산 오탄당 경로 (Pentose phosphate Pathway) (0) | 2022.07.31 |
| [생화학] 당분해 (Glycolysis) - 2 (0) | 2022.07.31 |
| [생화학] 당분해 (Glycolysis) - 1 (0) | 2022.07.31 |
| [생화학] 생물학적 산화-환원 반응 (Biological Oxidation-Reduction Reactions) (0) | 2022.07.30 |