지질은 물에 녹지 않는 불용성 생체 분자를 통칭한다. 지질은 매우 다양한 기능을 갖고 있기 때문에 지질 합성 경로는 모든 생물에게 있어 매우 중요하다. 지질 합성은 ATP를 소모하며, 환원형 전자 운반체로 NADPH를 필요로 한다.
우선 지방산의 생합성에 대해 논의한 후, 지방산으로 구성되는 트라이아실글리세롤, 인지질 합성 과정에 대해 알아볼 것이다. 이후 다른 지질인 콜레스테롤의 생합성에 대해 알아보도록 하자.

1. 개요

지방산은 에너지를 저장하기 때문에, 세포의 에너지 필요에 따라 지방산의 분해, 합성 경로는 반드시 구분되어야 한다. 지방산의 분해는 미토콘드리아 기질에서 일어나는 반면, 지방산의 합성은 동물 세포에서는 세포질, 식물세포에서는 엽록체에서 일어난다. 또한, 지방산 분해의 최종 산물은 아세틸-CoA지만, 지방산 합성의 전구체는 3탄소 화합물인 말로닐-CoA(Malonyl-CoA)다.
말로닐-CoA는 아세틸-CoA 카복실화효소(Acetyl-CoA carboxylase)에 의해 아세틸-CoA로부터 합성된다. 이 과정은 비가역적이며, 한 분자의 ATP를 소모한다. 형성된 말로닐-CoA는 지방산 합성 반응을 시작한다.
2. 지방산 합성
지방산 합성은 지방산 생성효소(Fatty acid synthase, FAS)에 의해 촉매된다. 지방산 생성효소는 크게 척추동물과 곰팡이의 FAS Ⅰ, 식물과 세균의 FAS Ⅱ 두 가지로 구분할 수 있다. FAS Ⅰ은 다기능 복합체이며, 지방산 합성에 필요한 7개의 활성자리가 하나의 단백질에 모여있다. FAS Ⅰ의 산물은 오직 팔미트산(Palmitic acid, 16:0)이다. FAS Ⅱ는 서로 떨어져 있는 여러 단백질 계로써, 불포화, 가지형 등 다양한 종류의 지방산 합성이 가능하다. 척추동물의 미토콘드리아도 FAS Ⅱ를 일부 가지고 있다. 이번 글에서는 팔미트산을 생성하는 FAS Ⅰ에 초점을 맞춰 지방산 합성을 논의할 것이다.

FAS Ⅰ은 두 개의 중요한 싸이올기(-SH)를 갖는다. 하나는 β-케토아실-ACP-생성효소(KS) 영역(회색)에 있는 Cys 잔기이고, 다른 하나는 아실 운반 단백질(Acyl carrier protein, ACP)의 -SH기이다. 지방산 합성의 최초 단계에서 말로닐-CoA는 ACP의 싸이올기와, 아세틸-CoA는 Cys 잔기와 결합해 있다. 그 후, 다음의 4단계를 거쳐 4탄소 지방산을 생성한다.
① 아세틸기의 축합

말로닐기의 말단 카복실기(초록색)가 떨어져나가며 아세틸기와 축합된다. 이 반응은 FAS Ⅰ의 KS 영역에 의해 촉매된다. 축합되어 형성된 아세토아세틸기는 ACP에 결합해있다. 이때 말로닐기의 카복실기를 어차피 제거할 것이라면, 왜 말로닐-CoA를 사용하는가? 이는 말로닐기가 아세틸기에 비해 더 불안정하고, 이를 이용해 자유 에너지 증가 반응인 지방산 합성을 보다 용이하게 하기 위함이다. 아세토아세틸기 생성 후 다음의 세 단계는 단순히 β 산화의 역과정이다.
② β 탄소의 환원

β 탄소의 케톤기는 NADPH에 의해 환원된다. 이 과정은 β-케토아실-ACP 환원효소(KR) 영역이 촉매한다. 이때 형성되는 중간체는 D형 이성질체인데, β 산화 중간체로 형성되는 L 이성질체와는 다르다는 것에 유의하라.
③ 단일결합의 탈수

한 분자의 물이 빠져나오며, α, β 탄소 간 이중결합이 형성된다. 이 과정은 β-하이드록시아실-ACP 수화효소(DH)에 의해 촉매된다.
④ α, β 탄소의 환원

최종적으로 이중결합이 환원되어 단일결합으로 바뀌고, 4탄소 아실인 뷰티릴-ACP(Butyryl-ACP)가 형성된다.
⑤ 아실-ACP의 전이
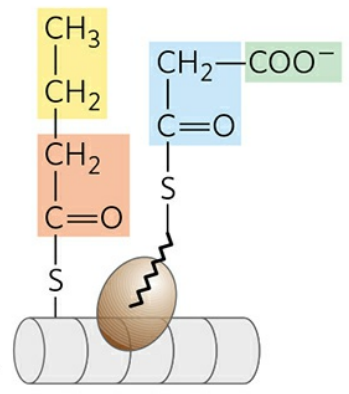
ACP와 결합해있던 뷰티릴기는 효소의 Cys 잔기로 전이된다. 이 과정은 말로닐/아세틸-CoA-ACP 전달효소(MAT)에 의해 촉매된다. 뷰티릴기는 지방한 합성의 최초 단계에서 아세틸기가 있던 위치로 옮겨진다. 이후, 새로운 말로닐기가 ACP에 결합해 위 일련의 단계들을 다시 수행한다.
위 과정이 7회 반복되면 16개 탄소로 이뤄진 팔미토일기가 ACP에 결합한 형태로 생성된다. 사슬 길이의 연장은 여기서 멈추고, 유리 팔미트산이 효소로부터 빠져나온다. 전체 효소 반응을 크게 두 부분으로 나눌 수 있다; 아세틸-CoA를 이용한 말로닐-CoA의 합성, FAS Ⅰ에 의한 축합.
$ 7Acetyl CoA + 7CO_2 + 7ATP \rightarrow 7Malonyl CoA + 7ADP + 7P_i $
$ Acetyl CoA + 7Malonyl CoA + 14NADPH + 14H^+ \rightarrow Palmitic acid + 7CO_2 + 8CoA + 14NADP^+ + 6H_2O $
두 반응을 합치면, 1분자의 팔미트산을 합성하기 위해 8개의 아세틸-CoA, 7개의 ATP, 14개의 NADPH가 소모됨을 알 수 있다. 아세틸-CoA가 세포질로 나오는 과정에서 추가적으로 2분자의 ATP를 더 소모하므로, 하나의 팔미트산을 합성할 때 소모되는 전체 ATP는 23개가 된다.
3. NADPH의 생성
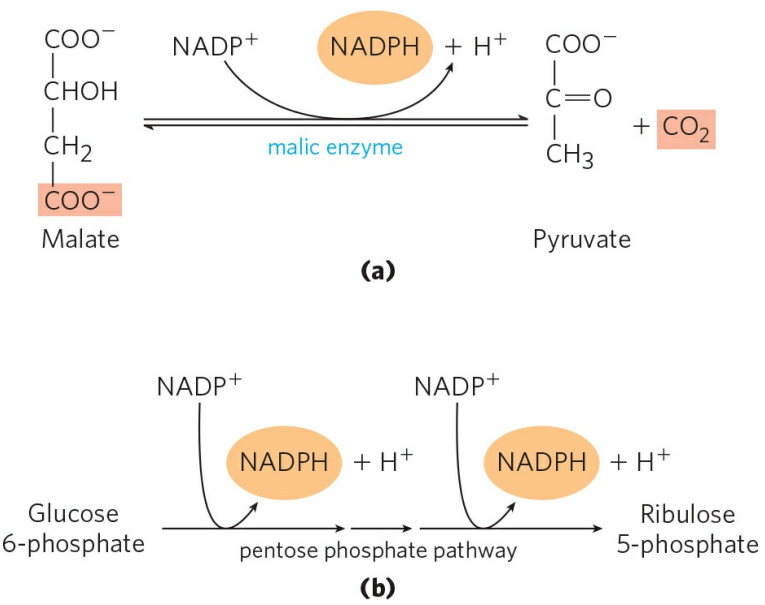
동물의 간세포와 지방세포에서 NADPH는 주로 앞서 배운 인산 오탄당 경로와 말산 효소(Malic enzyme)에 의해 만들어진다. 말산 효소는 말산을 탈카복실화하며, 생성된 피루브산은 미토콘드리아 내부로 수송된다. 식물세포에서 NADPH는 명반응에 의해 생성된다. 때문에 식물세포의 지방산 합성은 엽록체의 버팀질(Stroma)에서 일어난다.
4. 아세틸-CoA의 수송
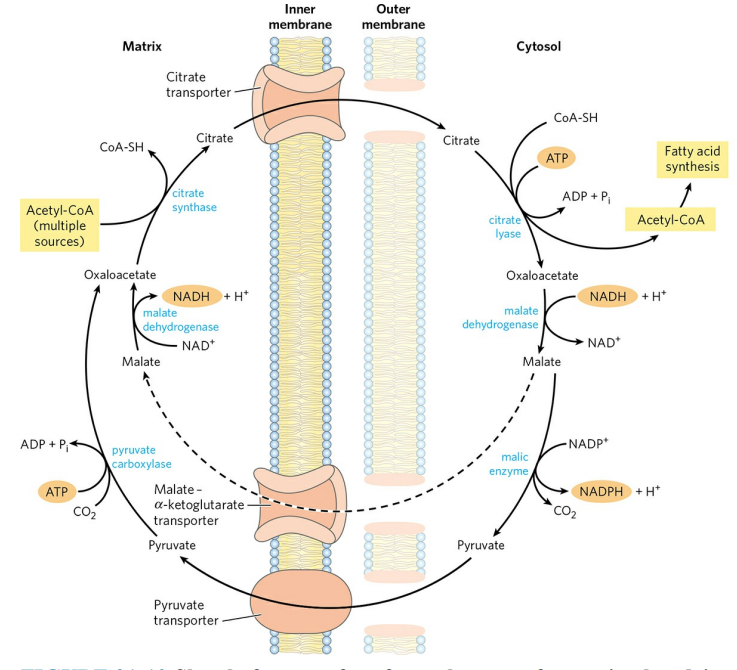
아세틸-CoA는 미토콘드리아 내막을 통과할 수 없다. 때문에 지방산 합성에 요구되는 아세틸기는 간접적인 통로를 통해 미토콘드리아를 빠져나온다. 우선, 아세틸-CoA는 옥살아세트산과 반응해 시트르산이 된다. 이 반응은 시트르산 회로 반응이다. 이후, 시트르산은 시트르산 수송체를 통해 세포질로 빠져나온다. 세포질에서 시트르산 분해효소(Citrate lyase)는 ATP를 사용해 시트르산을 분해하고, 옥살아세트산과 아세틸-CoA를 생성한다.
옥살아세트산은 말산으로 전환된다. 말산은 바로 미토콘드리아 내부로 유입될 수 있지만, 이 과정에서 생성된 말산은 대개 말산 효소의 작용을 받아 피루브산이 되며 NADPH를 생성한다. 지방산 합성에 필요한 NADPH의 절반 정도가 이 과정에서 만들어진다. 이후, 피루브산은 피루브산 수송체에 의해 미토콘드리아 내부로 유입되고, 피루브산 카복실화효소에 의해 옥살아세트산으로 전환된다. 이 과정에서 1분자의 ATP를 더 소모한다. 즉, 이 통로를 이용하면 1분자의 아세틸-CoA당 2분자의 ATP를 소모하며 1분자의 NADPH를 만드는 셈이 된다.
5. 지방산 합성의 조절
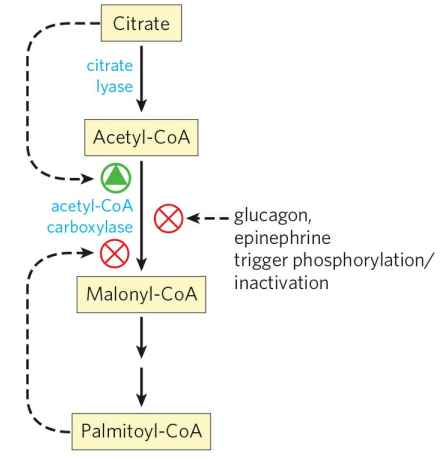
지방산 합성의 속도 제한 단계는 아세틸-CoA 탈카복실화효소 반응이다. 척추동물에서 지방산 합성의 주된 산물인 팔미토일-CoA는 이 효소를 억제하며, 과량의 시트르산(고에너지 신호)는 이 효소를 활성화하는 입체다른자리 조절자다. 글루카곤, 에피네프린은 아세틸-CoA 탈카복실화효소를 인산화시켜 불활성형으로 바꿔 지방산 합성을 저해한다. 식물세포에서 이 효소는 빛이 많을 때 높아지는 pH, [Mg+]에 의헤 활성화된다. 세균에서 지방산은 에너지 저장 분자가 아니므로, 합성 조절은 오직 세포 증식과 막의 형성의 필요에 의해 조절된다.
'생화학 > 생화학 (하)' 카테고리의 다른 글
| [생화학] 광합성 (Photosynthesis) - 1 (0) | 2022.09.05 |
|---|---|
| [생화학] 산화인산화 (Oxidative Phosphorylation) - 3 (0) | 2022.09.05 |
| [생화학] 산화인산화 (Oxidative Phosphorylation) - 2 (0) | 2022.09.03 |
| [생화학] 산화인산화 (Oxidative Phosphorylation) - 1 (0) | 2022.09.01 |
| [생화학] 아미노산 분해 경로 (Pathways of Amino Acid Degradation) (0) | 2022.09.01 |