지금까지 아미노산의 공통 아미노기 제거와 배설에 대한 논의를 했다면, 이번 글에서는 각각의 아미노산이 정확히 어떤 경로를 거쳐 이화되는지에 대해 알아보도록 하자.
아미노산의 이화는 체내 에너지 생산의 10~15% 정도만을 담당한다. 20가지 아미노산의 이화 경로는 6개의 주요 산물로 수렴하고, 산물들은 모두 시트르산 회로로 유입된다. 이번 글에서는 아미노산의 이화 경로 중 특히 중요한 몇 가지 경로와 아미노산 이화와 관련된 질병에 대해 중점적으로 다뤄보도록 하자. 다양한 아미노산에 대한 이야기를 하고 있으므로 20종 아미노산의 구조를 알아두는 것이 좋다.

1. 아미노산 분해 개요
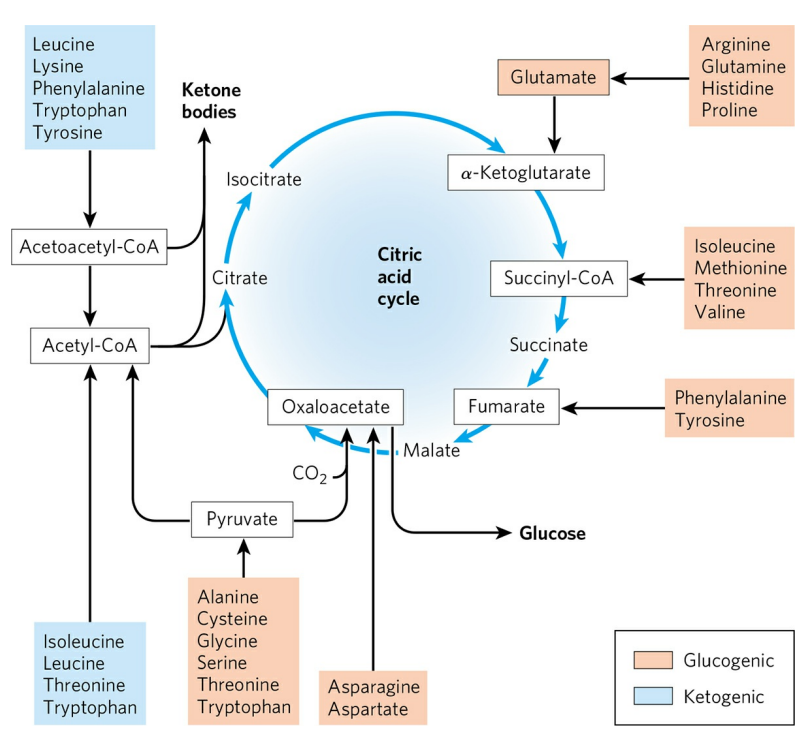
거의 대부분의 아미노산 분해 경로는 6개의 주요 산물(아세틸-CoA, 피루브산, α-케토글루타르산, 석시닐-CoA, 퓨마르산, 옥살아세트산)로 수렴한다. 6개의 산물은 모두 시트르산 회로로 유입될 수 있다. 거대한 아미노산의 경우 각각의 조각이 다른 경로를 따르기도 하고, 하나의 아미노산이 두 가지 이상의 이화 경로로 대사되기도 한다.
아미노산은 크게 케톤생성(Ketogenic) 아미노산과 포도당생성(Glucogenic) 아미노산으로 나눌 수 있다. 케톤생성 아미노산은 이화의 최종 산물이 아세토아세틸-CoA, 아세틸-CoA이며, 필요에 따라 시트르산 회로로 유입되지 않고 간에서 케톤체 형성에 쓰일 수 있다. 당뇨병 환자의 간에서는 케톤 생성 아미노산의 이화 경로로부터 많은 양의 케톤체가 생성되어 케토산증에 기여한다. 두 아미노산간의 구분은 명확하지 않다. 트립토판, 페닐알라닌, 타이로신, 트레오닌, 아이소류신은 케톤과 포도당 둘 다 생성할 수 있다.
2. 아미노산 이화 경로의 보조인자
아미노산 이화 경로 중 아미노기를 제거하는 트랜스아미네이션 반응은 보조인자로 PLP를 이용한다고 언급했었다. 이와 유사하게, 아미노기 이화 경로 중 일탄소 전이(One-carbon transfer) 반응에 참여하는 3가지 보조인자가 있다; 바이오틴, 사수소폴산(Tetrahydrofolate), S-아데노실메싸이오닌(S-Adenosylmethionine)
1) 바이오틴(Biotin)

바이오틴은 다양한 효소에서 사용되는 보조인자다. 바이오틴은 일탄소 단위 중 이산화탄소(중탄산염)를 카복실기의 형태로 전달하는 기능을 한다. 위 그림에서처럼 바이오틴 고리의 질소 원자가 이산화탄소와 결합해 효소의 기질로 카복실기를 전달해준다.
2) 사수소폴산(Tetrahydrofolate, H4 Folate)

사수소폴산은 6-메틸테린, p-아미노벤조산, 글루탐산으로 구성되는 복잡한 분자다. 산화형은 비타민 B6다. 사수소폴산은 메틸기(-CH3), 포르밀기(-CH=O), 하이드록시메틸렌기(-CH2OH) 등 다양한 산화 상태의 일탄소 단위를 전달하는 데 이용된다. 사수소폴산의 중요한 작용 부위는 6-메틸테린의 5번과 10번 질소(하늘색으로 표시) 원자다.
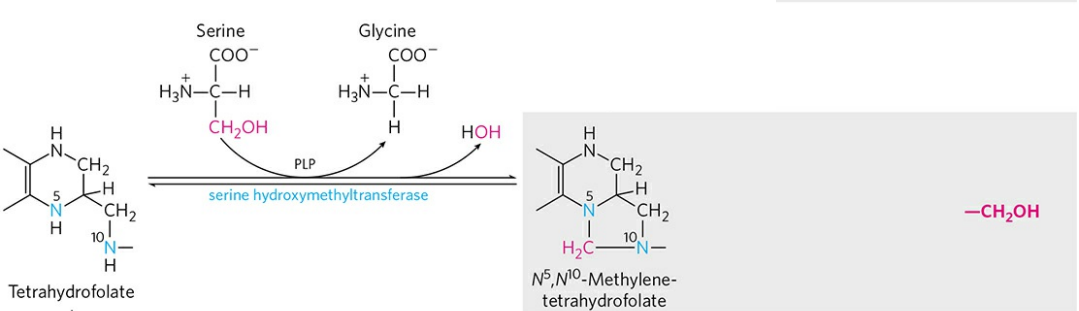
사수소폴산의 일탄소 단위를 전달하는 주요 공급원은 세린이다. 세린은 자신의 하이드록시메틸렌기를 사수소폴산에 전달해 글라이신으로 전환되고, 사수소폴산의 5, 10번 질소는 전달받은 메틸렌기와 고리 구조를 이뤄 N5, N10-메틸렌사수소폴산을 형성한다. 사수소폴산의 다른 일탄소 단위 운반은 나중에 다시 다루도록 하자.
3) S-아데노실메싸이오닌(S-Adenosylmethionine, adoMet)
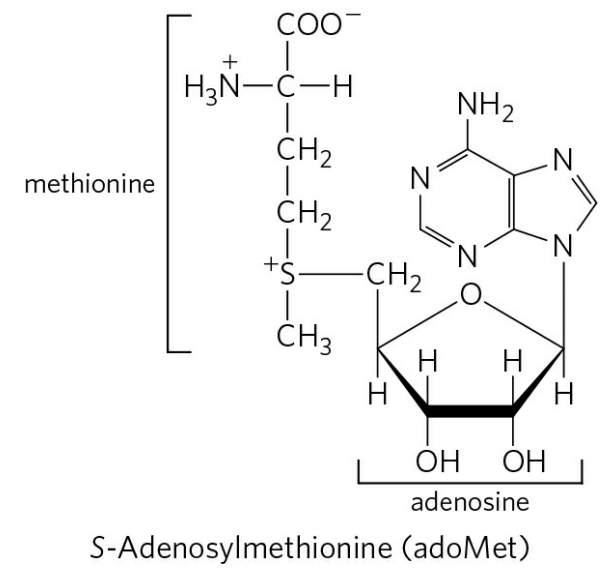
S-아데노실메싸이오닌은 아데노신에 메싸이오닌이 결합한 화합물이며, 메싸이오닌 아데노실 전달효소에 의해 합성된다. S-아데노실메싸이오닌은 메틸기(-CH3)를 전달하는 데 특화되어 있는데, 전달 과정에서 하나의 메틸 회로(Methyl cycle)를 형성한다.
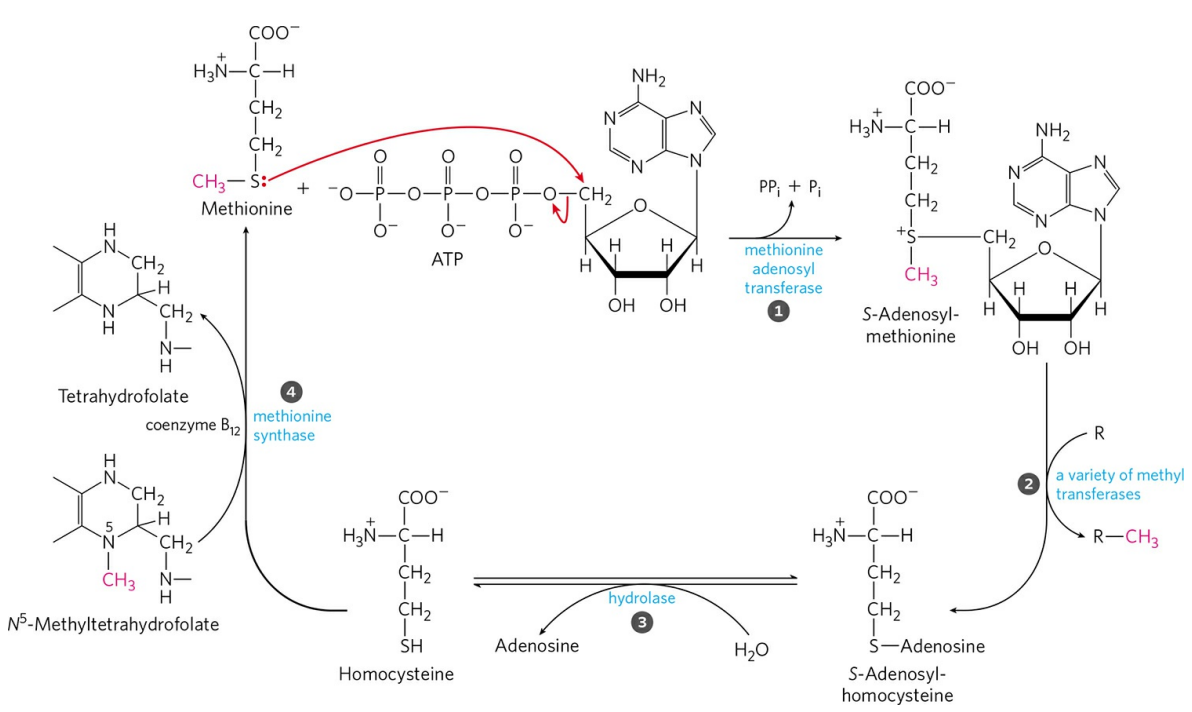
메싸이오닌은 ATP의 아데노신과 결합해 S-아데노실메싸이오닌을 형성한다(①). 이 과정은 ATP에서 삼인산을 분리시키는 드문 반응들 중 하나다. S-아데노실메싸이오닌의 메틸기(붉은색으로 표시됨)는 다양한 메틸기전달효소에 의해 전달된다(②). 이후, 아데노신은 가수분해되어 떨어져나오고, 메싸이오닌에서 메틸기가 제거된 호모시스테인(Homocysteine)이 남는다(③). 호모시스테인은 메틸기 주개(위 그림에서는 메틸사수소폴산)로부터 메틸기를 전달받아 메싸이오닌으로 재생된다(④).
3. 아미노산 분해
1) 피루브산 생성 : 알라닌, 트립토판, 시스테인, 세린, 글라이신, 트레오닌
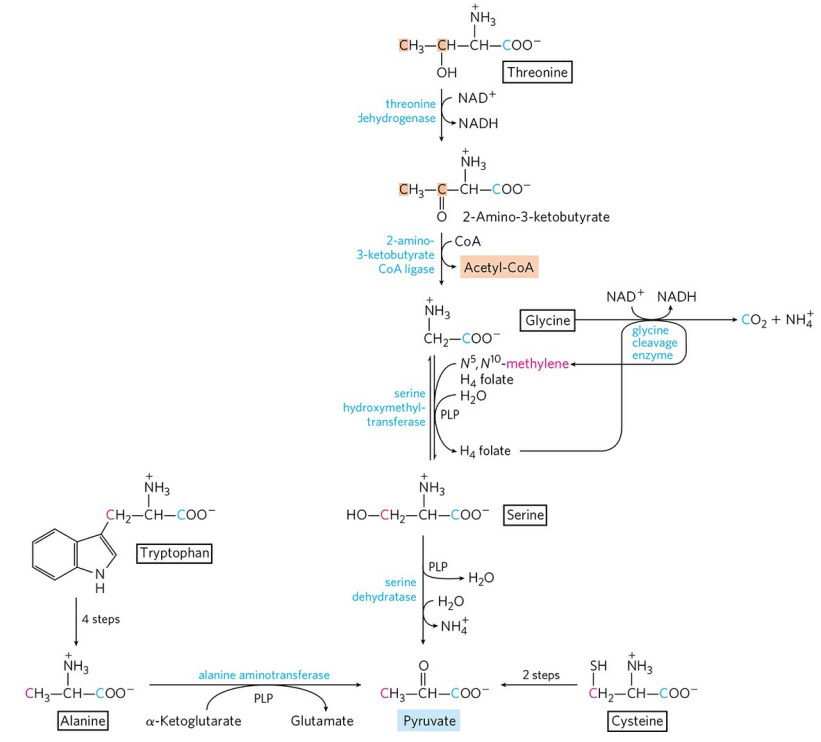
알라닌은 앞서 언급했듯 피루브산으로의 전환이 자유롭다. 트립토판은 고리 제거 후 알라닌으로 전환된다. 고리는 다른 경로를 따라 분해된다. 시스테인은 황 원자 제거 후 아미노기가 제거되어 피루브산으로 전환된다. 트레오닌, 글라이신은 몇 단계를 거쳐 세린이 되고, 세린은 세린 탈수효소에 의해 피루브산으로 전환된다. 세린 탈수효소는 PLP를 필요로 한다. 세린을 경유하는 트레오닌 이화 경로는 비교적 부수적인 경로이고, 주요 경로는 석시닐-CoA로 향하는 경로이다.
글라이신은 세린이 되는 과정 이외에 두 가지 이화 경로를 추가로 갖는다. 글라이신 절단효소(Glycine cleavage enzyme)에 의한 반응은 위 그림에 표시되어 있다. 글라이신의 카복실기는 이산화탄소의 형태로 소실되고, 암모늄 이온은 배설되며, α 탄소는 메틸렌기가 되어 사수소폴산에 전달된다. 이 경로는 동물에서 특히 중요해, 글라이신 절단효소 활성에 결함이 있으면 혈중 글라이신 농도가 높아지는데 이는 매우 유독하다.
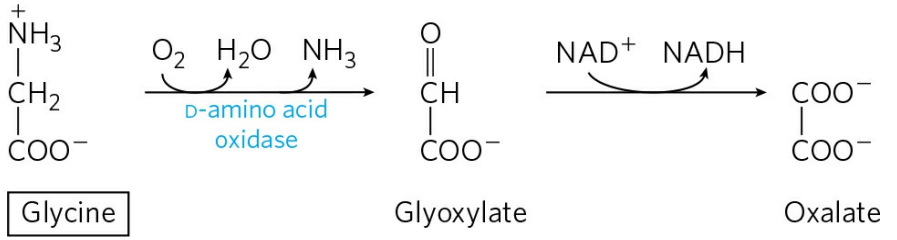
글라이신 이화의 세 번째 경로는 D-아미노산 산화효소에 의해 촉매된다. D-아미노산 산화효소는 세균 감염, 구운 식품 섭취 등으로 인한 D-아미노산의 해독을 담당한다. 이 반응에 의해 생성된 옥살산염은 칼슘과 결합해 신장결석의 형태로 배설된다.
2) 아세틸-CoA 생성 : 트립토판, 라이신, 페닐알라닌, 타이로신, 류신, 아이소류신, 트레오닌
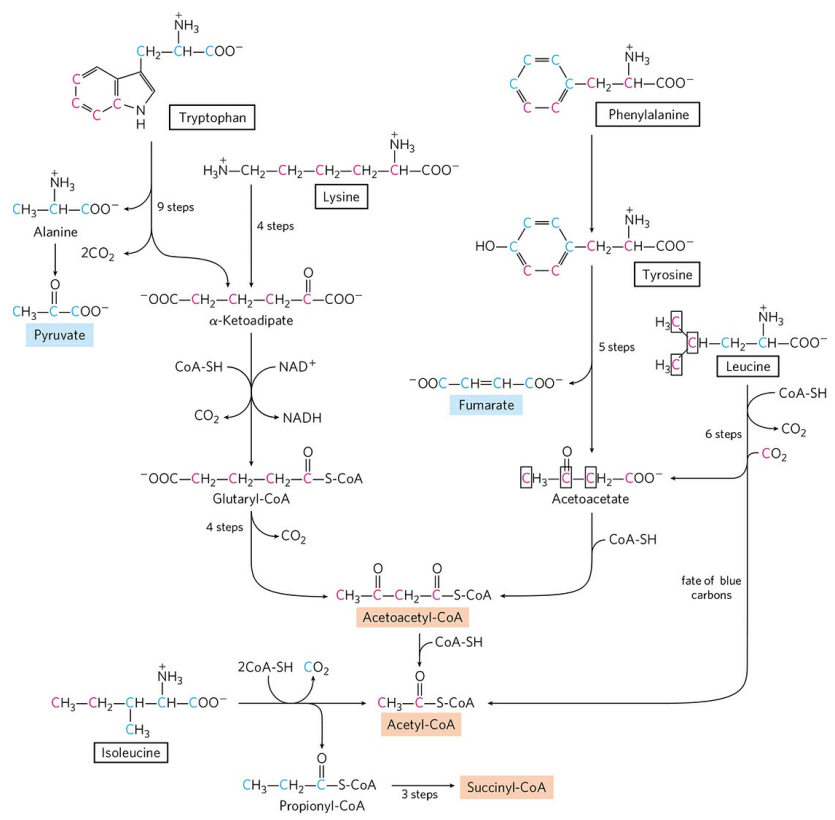
트립토판의 일부분(파란색으로 표시됨)은 앞서 언급했듯 알라닌을 거쳐 피루브산을 생성한다. 트립토판 나머지 고리의 일부분은 복잡한 과정을 거쳐 아세토아세틸-CoA를 생성한다. 라이신 또한 트립토판과 비슷한 경로를 거쳐 아세토아세틸-CoA를 생성한다. 류신, 아이소류신은 탄소 골격의 일부는 아세틸-CoA, 나머지 일부는 각각 아세토아세틸-CoA와 석시닐-CoA를 생성한다.
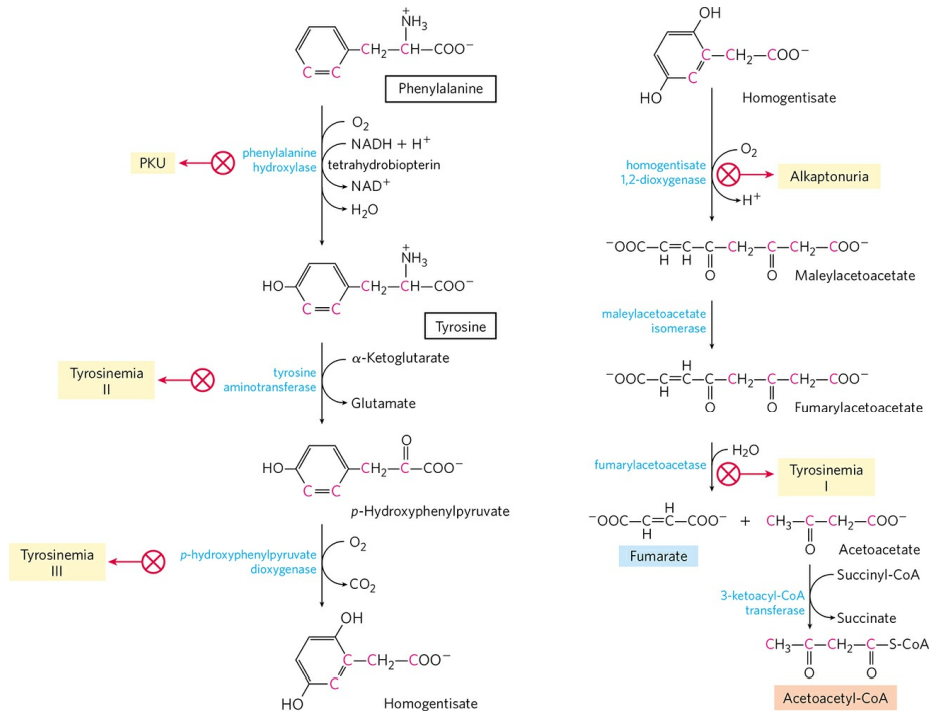
페닐알라닌 이화 경로 효소의 유전적 결함은 다양한 유전병을 일으킨다. 페닐알라닌은 우선 타이로신으로 수산화될 필요가 있다. 이 반응을 촉매하는 페닐알라닌 수산화효소(Phenylalanine hydroxylase)의 결함은 유명한 유전병인 페닐케톤뇨증(Phenylketonuria, PKU)을 유발한다. 이후 타이로신 이화 단계를 촉매하는 효소의 유전적 결함은 여러 종류의 타이로신혈증(Tyrosinemia)을 유발한다. 타이로신의 이화 과정 중 고리 구조를 끊는 호모젠티스산 이산소화효소(Homogentissate dioxygenase)의 결함은 알캅톤뇨증(Alkaptonuria)을 유발한다.
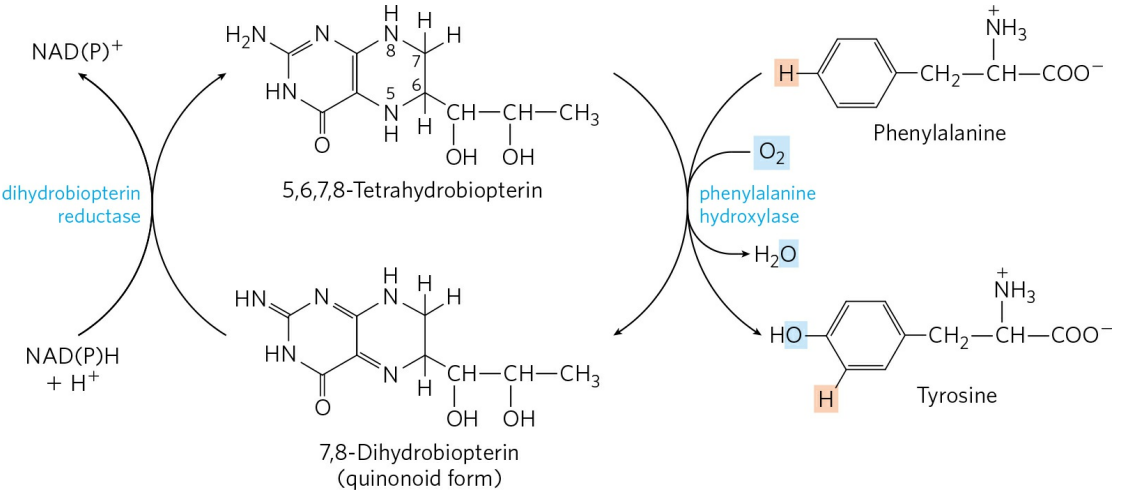
페닐알라닌 수산화효소는 보조인자로 사수소바이오테린을 필요로한다. 사수소바이오테린은 효소 작용에 소모되면 이수소바이오테린으로 산화되고, 이는 이수소바이오테린 환원효소에 의해 다시 사수소바이오테린으로 재생된다. 이수소바이오테린 환원효소의 결함 또한 페닐알라닌 수산화효소의 작용을 못하게 하므로 페닐케톤뇨증을 유발한다.
3) α-케토글루타르산의 생성 : 프롤린, 글루탐산, 글루타민, 아르지닌, 히스티딘
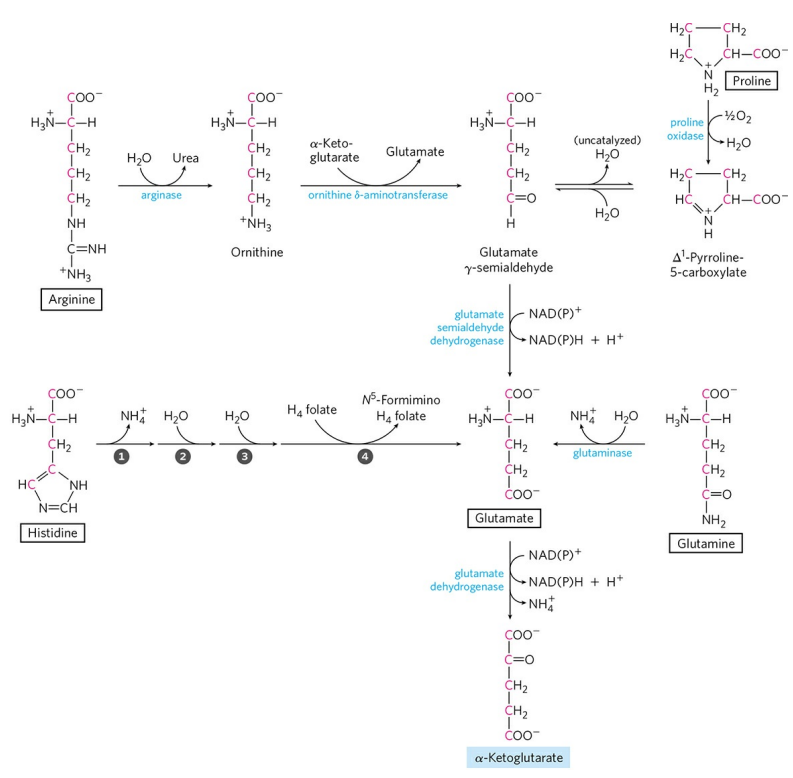
프롤린, 글루타민, 아르지닌, 히스티딘은 각각의 경로를 통해 글루탐산으로 수렴되는 이화 경로를 갖는다. 글루탐산은 앞서 언급했던 글루탐산 탈수소효소에 의해 α-케토글루타르산으로 탈아미노화 된다.
4) 석시닐-CoA 생성 : 메싸이오닌, 아이소류신, 트레오닌, 발린
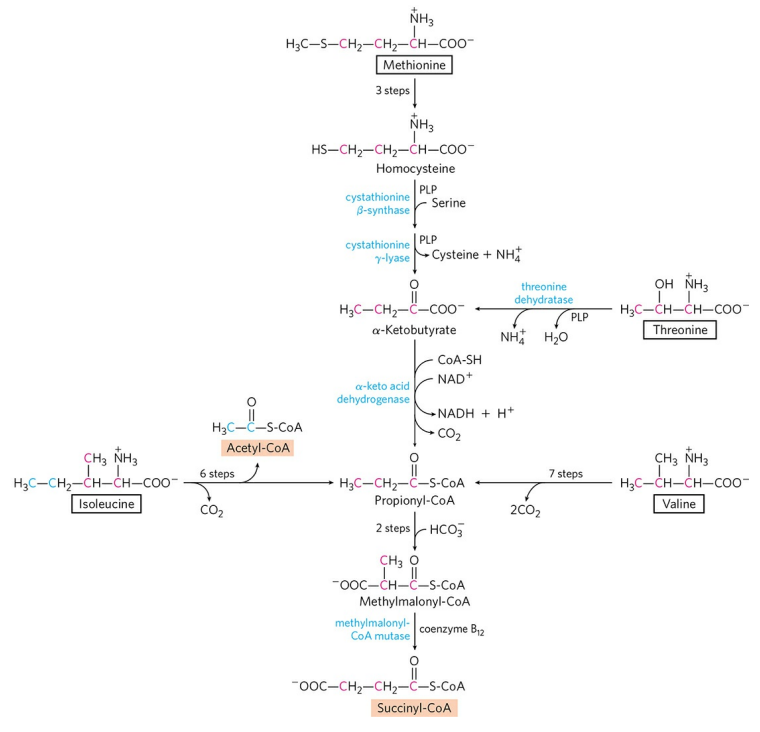
메싸이오닌, 아이소류신, 트레오닌, 발린은 프로피오닐-CoA를 거쳐 석시닐-CoA를 생성한다. 프로피오닐-CoA가 석시닐-CoA로 전환되는 반응은 홀수 지방산의 산화에서 다룬 적이 있다. 아이소류신의 2개 탄소 원자는 아세틸-CoA로 빠져나간다는 점에 유의하라.
5) 옥살아세트산의 생성 : 아스파라진, 아스파트산
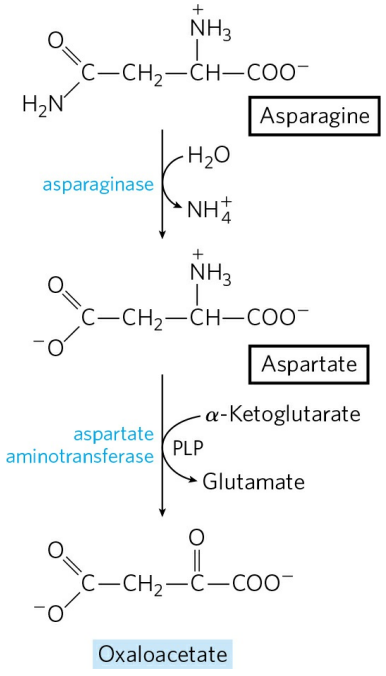
아스파라진은 아스파라진분해효소(Asparaginase)에 의해 쉽게 아스파트산으로 전환된다. 아스파트산이 옥살아세트산으로 전환되는 과정은 요소 회로의 준비기에서 언급한 적이 있다.
6) 추가 경로 : 가지사슬 아미노산의 이화
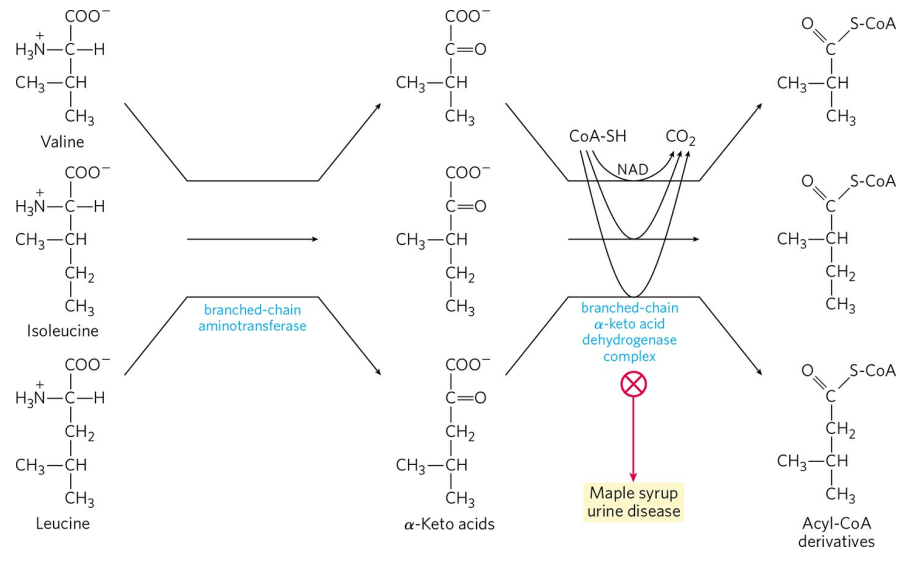
대부분의 아미노산 이화 반응은 간에서 일어나지만, 가지사슬 아미노산 3종류(발린, 아이소류신, 류신)는 근육, 지방, 신장, 뇌에서 에너지원으로 이용되기도 한다. 이들은 간외 조직에 존재하는 가지사슬 아미노기 전달효소에 의해 각각의 케토산으로 전환되고, 이후 가지사슬 α-케토산 탈수소효소 복합체에 의해 CoA가 붙어 아실-CoA를 형성한다. α-케토산 탈수소효소 복합체는 피루브산 탈수소효소 복합체와 본질적으로 동일한 반응 기전을 갖는다. α-케토산 탈수소효소 복합체의 유전적 결함은 α-케토산의 축적으로 인한 단풍시럽뇨병(Maple syrup urine disease)을 유발한다. 이 질병은 발린, 아이소류신, 류신의 섭취를 최소한으로 제한하는 엄격한 식이 조절을 필요로 한다.
지금까지 20종 아미노산의 대략적인 이화 경로에 대해 알아보았다. 세세한 이화 경로를 전부 알 필요 없이, 이화 반응에 쓰이는 보조인자와 다양한 경로를 갖는 아미노산(글라이신 등), 특이한 유전병(PKU, 알캅톤뇨증, 단풍시럽뇨병 등)과 관련된 이화 경로에 대해서만 중점적으로 알고 있으면 된다. 다음 글에서는 탄수화물, 지방, 단백질 이화 경로의 궁극적 목표인 산화인산화(Oxidative phosphorylation)에 대해 다뤄보도록 하자.
'생화학 > 생화학 (하)' 카테고리의 다른 글
| [생화학] 산화인산화 (Oxidative Phosphorylation) - 2 (0) | 2022.09.03 |
|---|---|
| [생화학] 산화인산화 (Oxidative Phosphorylation) - 1 (0) | 2022.09.01 |
| [생화학] 요소 회로 (Urea cycle) (0) | 2022.09.01 |
| [생화학] 아미노기 대사 (Metabolism of Amino Groups) (0) | 2022.08.21 |
| [생화학] 케톤체 (Ketone Bodies) (0) | 2022.08.17 |