앞선 글에서 아실-CoA가 카니틴 왕복 통로를 통해 미토콘드리아 내부로 유입되는 과정까지 알아보았다. 이번 글에서는 미토콘드리아 내부로 유입된 지방산이 어떻게 산화되는지에 대해 알아보도록 하자.

1. 지방산의 산화 개요
미토콘드리아 내부로 유입된 아실-CoA의 공통 구조는 다음과 같다.

지방산의 산화는 아실-CoA의 탄소 2개가 아세틸-CoA의 형태로 떨어져 나가며 진행된다. 대략적인 반응 그림은 다음과 같다.

아실-CoA의 β 탄소가 산회되어 카보닐기를 형성하면, β 탄소와 α 탄소 단일결합의 분해가 일어날 수 있다. 분해된 β 탄소에는 다시 CoA가 결합해 위 반응을 다시 수행한다. 이 반응은 β 탄소가 산화므로 β 산화(β Oxidation)라 불리고, β 산화는 지방산 이화의 주요 경로이다. β 산화를 통해 생성된 아세틸-CoA는 시트르산 회로로 유입된다.
이번 글에서는 우선 일반적인 포화 지방산의 β 산화 대해 다루고, 그다음에 불포화 지방산, 홀수 지방산 등의 β 산화에 대해 이야기하도록 하자. 주요하지 않은 경로로 ω, α 산화도 존재하는데, 이들 경로에 대해서는 밑에서 간략하게 다뤄보도록 하자.
2. β 산화
지방산에서 탄소들를 연결하는 에틸렌 결합(-CH2-CH2-)은 매우 안정적이다. β 산화는 이러한 에틸렌 결합을 불안정하게 만든 후 절단하는 매우 정밀한 기전이다. β 산화의 효소는 지방산 길이에 특이적이다. 12개 이상의 탄소를 갖는 지방산은 미토콘드리아 내막에 결합한 삼기능 단백질(Trifunctional protein, TFP)에 의해 분해된다. 이들은 반응이 일어나는 동안 기질 통로화를 통해 지방산 사슬을 잡아둔다. 탄소 수가 12개 이하로 떨어지면, 그 이후의 β 산화는 미토콘드리아 바탕질에 존재하는 효소에 의해 촉매된다.
1) 짝수, 포화 지방산
β 산화는 쉽게 말해 β 탄소의 메틸렌기(-CH2-)가 카보닐기(-(C=O)-)로 산화되는 과정이다. 이 반응의 자세한 중간 과정은 다음과 같다.
① α, β 탄소의 탈수소화

α, β 탄소가 모두 탈수소화되며 탄소 간 이중결합을 형성한다. 이 반응은 아실-CoA 탈수소효소(Acyl-CoA dehydrogenase)에 의해 촉매된다. 이 효소는 지방산의 길이에 따라 다양한 동종효소를 갖는다. 이 동종효소들은 모두 FAD를 조효소로 갖는 플라보단백질(Flavoprotein)이다.
② 이중결합의 수화
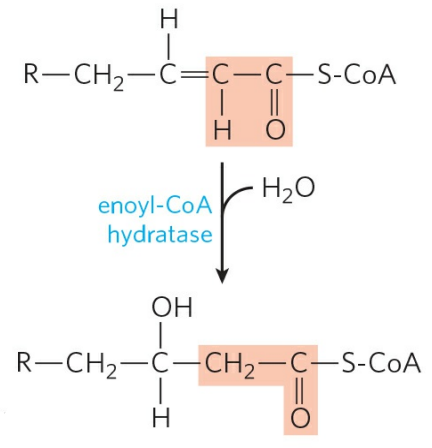
탄소-탄소 이중결합은 에노일-CoA 수화효소(Enoyl-CoA hydratase)에 의해 수화된다. 이 반응은 결과적으로 β의 수소 하나가 하이드록실기로 치환된 β-하이드록시아실-CoA(β-Hydroxyacyl-CoA)를 형성하는 셈이 된다. 여기서 물 한 분자가 소비된다.
③ β 탄소의 산화

수화된 β 탄소는 다시 β-하이드록시아실-CoA 탈수소효소(β-Hydroxyacyl-CoA dehydogenase)에 의해 탈수소화되고, β 탄소에 케톤기가 형성된다. β-하이드록시아실-CoA 탈수소효소는 NAD+를 조효소로 갖는다.
④ 아세틸-CoA의 형성

아실-CoA 아세틸기전달효소(Acyl-CoA acetyltransferase)가 α, β 탄소 간 단일결합을 CoA로 절단한다. 이 절단은 CoA의 싸이올기(-SH)을 이용한 싸이올 분해(Thiolysis) 작용이다. 분해된 α 탄소 부분은 아세틸-CoA로 떨어져나가고, β 탄소에는 CoA가 결합해 원래보다 탄소가 2개 적어진 새로운 아실-CoA를 형성한다. 새로운 아실-CoA는 다시 β 산화를 반복해서 수행한다.

β 산화의 기전은 시트르산 회로의 석신산 → 옥살아세트산 기전과 정확히 같다. 석신산은 FAD에 의해 탈수소화되어 퓨마르산을 형성하고, 퓨마르산은 수화되어 말산을 형성하며, 말산은 다시 NAD+에 의해 탈수소화되며 최종적으로 옥살아세트산을 형성한다. 이는 유전자 중복(Gene dupilcation)에 의한 기전 보전성의 대표적인 예시다.

β 산화는 그 과정 자체로 FADH2, NADH 한분자씩을 생성하고, 분해 산물인 아세틸-CoA 또한 시트르산 회로를 거치며 FADH2, NADH를 생성한다. 생성된 FADH2, NADH는 전자전달계를 거쳐 ATP를 합성한다. 시트르산 회로, 전자전달계는 모두 유산소(Aerobic) 환경을 필요로 한다. 즉, 지방산 산화는 유산소 환경에서만 에너지를 생성할 수 있다.
18개 탄소로 이뤄진 팔미토일-CoA는 완전 산화될 경우 108 분자의 ATP가 생성된다. 미토콘드리아 밖에서 팔미트산이 팔미토일-CoA로 전환될 때 ATP(AMP를 이용하므로 ATP 2개 분량의 에너지)를 사용함을 생각해보면, 팔미트산 1분자는 106 분자의 ATP를 생성하는 셈이 된다. 이는 표준 상태에서 33%, 세포 내 상태에서 60% 이상의 에너지 수율을 갖는다.
2) 불포화 지방산
불포화 지방산은 이중결합이 하나 이상 포함된 지방산이다. 이중결합은 시스와 트랜스, 두 가지 형태를 갖는다.

이중결합 자체는 β 산화의 ① 단계에서도 형성되므로 문제가 없다. 다만 자연 불포화지방산의 대부분은 시스 지방산인 반면, β 산화 중 형성되는 이중결합은 트랜스 형태이다. 때문에 β 산화를 수행하기 위해 시스 이중결합이 트랜스 이중결합으로 전환될 필요가 있다.
불포화 지방산의 단일 결합 부분은 β 산화를 거친다. 그 후, 더 이상 β 산화가 진행될 수 없을 때 불포화 지방산의 이중결합은 보통 γ 탄소와 β 탄소 사이에 위치한다.

이러한 시스 결합은 Δ3, Δ2-에노일-CoA 이성화효소(Δ3, Δ2-enoyl-CoA isomerase)에 의해 트랜스 결합으로 전환된다.

결합의 형태가 전환될 때 이중결합의 위치 또한 바뀌므로, 이후 바로 β 산화의 ② 과정이 진행될 수 있다.
다음은 흔한 단일 불포화 지방산 중 하나인 올레산(Oleate)의 산화 과정이다.

다불포화 지방산(Polyunsaturated fatty acid)의 경우 조금 더 복잡한 과정을 거친다. 일반적인 다불포화 지방산에서 두 개의 이중결합은 다음과 같이 위치한다.

γ-β 이중결합은 위에서처럼 이성화 효소에 의해 트랜스 결합으로 전환되고, β 산화가 진행된다.

남은 불포화 사슬은 다시 β 산화의 ① 과정을 거치고, δ-γ 이중결합이 형성된다. ζ-ε, δ-γ 두 이중결합은 NADPH에 의해 환원되고 동시에 축합되며 ε-δ 이중결합을 형성한다.

ε-δ 이중결합은 에노일-CoA 이성화효소(Enoyl-CoA isomerase)에 의해 δ-γ 이중결합으로 위치가 바뀐다.

δ-γ 이중결합을 갖는 이 반응의 산물은 β 산화의 ② 반응을 진행할 수 있게 된다. 이후 β 산화를 거쳐 완전히 산화된다.
다음은 흔한 다불포화 지방산인 리놀레산(Linoleic acid)의 산화 과정이다.

정리하면, 불포화 지방산은 β 산화에 이용되는 효소 외에 이성질화 효소 및 환원효소를 추가적으로 필요로 한다.
3) 홀수 지방산
홀수 지방산은 많은 식물과 일부 해양생물이 함유하고 있다. 특히 빵과 시리얼에는 3탄소 지방산인 프로피온산(Propionate)이 소량 들어있다. 홀수 지방산은 β 산화를 거쳐 2n개의 탄소를 아세틸-CoA의 형태로 분해되고, 최종적으로 프로피온산의 CoA 형인 프로피오닐-CoA(Propionyl-CoA)가 한 분자 남게 된다.
프로피오닐-CoA는 프로피오닐-CoA 카복실화효소(Propionyl-CoA carboxylase)에 의해 카복실화되어 4탄소 화합물인 D-메틸말로닐-CoA(D-Methylmalonyl-CoA)를 형성한다.

D-메틸말로닐-CoA는 메틸말로닐-CoA 에피머화효소(Methylmalonyl-CoA epimerase)에 의해 L-메틸말로닐-CoA로 전환된다. L-메틸말로닐-CoA는 다시 메틸말로닐-CoA 변위효소에 의해 석시닐-CoA로 전환되고, 석시닐-CoA는 시트르산 회로로 유입된다. 석시닐-CoA로의 전환은 비타민 B12로부터 유래된 보조효소 B12(Coenzyme B12)를 필요로 한다.

3. β 산화의 조절
β 산화, 지방산의 산화는 귀중한 연료를 소비하므로, 반드시 고에너지를 필요로 하는 환경에서만 산화되어야 한다. 지방산 산화의 조절 단계는 카니틴 왕복통로이므로, 지방산의 산화는 아실-CoA가 미토콘드리아 내부로 유입되는 카니틴 왕복통로에서 조절된다.
에너지원으로 바로 이용할 수 있는 포도당의 혈중 농도가 높아지면 체내에서는 인슐린이 분비된다. 인슐린은 아세틸-CoA 카복실화효소(Acetyl-CoA carboxylase, ACC)를 탈인산화해 활성화시킨다. 활성화된 ACC는 아세틸-CoA를 카복실화해 말로닐-CoA(Malonyl-CoA)를 생성한다. 말로닐-CoA는 지방산 합성의 전구체인데, 때문에 말로닐-CoA는 카니틴 아실기전달효소 1을 억제해 지방산이 미토콘드리아 내부로 들어가는 것을 막고, 지방산 합성을 촉진한다.

반면, 포도당이 유입되지 않는 공복 상태에서는 혈당이 떨어져 글루카곤이 분비된다. 글루카곤은 ACC를 인산화해 불활성화시킨다. 이 과정은 AMP도 촉진한다. ACC가 불활성화되면 말로닐-CoA의 농도가 떨어지고, 지방산은 카니틴 아실기전달효소에 의해 미토콘드리아 내부로 유입되어 산화될 수 있다. 글루카곤은 또한 지방산의 동원을 촉발하기도 했던 것을 기억하는가? 글루카곤은 포도당신생성을 촉진하며, 동시에 포도당 대신 지방을 에너지원으로 이용하도록 작용한다.

β 산화의 과정 또한 조절받는다. [NADH]가 높아지면 β 산화 ③ 반응의 효소를 억제하고, 아세틸-CoA는 β 산화 ④ 반응의 효소를 억제한다.
장기적인 관점에서, 지방산의 산화는 전사인자에 의해 조절받기도 한다. PPAR 패밀리 중 PPARα는 지방산 운반체, 카니틴 아실기 전달효소 1과 2, 긴 사슬 지방산 아실-CoA 탈수소효소 등의 유전자 체계를 작동시킨다. 이들은 오랜 굶주림(Starvation)과 같은 상태에서 촉발된다. 글루카곤 또한 전사인자인 CREB를 조절해 지방 대사에 관련된 특정 유전자 발현을 조절하기도 한다.
4. 퍼옥시좀에서의 β 산화
퍼옥시좀(Peroxisome, 과산화소체라고도 불림)은 진핵세포의 세포질 내에 존재하는 세포 소기관으로, 긴사슬 지방산, 독성 물질 등을 분해하는 역할을 한다. 동물세포에서 β 산화는 대부분 미토콘드리아에서 일어나지만, 식물세포에서 β 산화는 대부분 퍼옥시좀에서 일어난다.
두 세포 소기관에서 일어나는 β 산화 과정은 거의 유사하지만, 두 가지 차이점이 있다. 첫째로, 퍼옥시좀의 β 산화로 생성되는 FADH2는 전자전달계로 가지 않고 과산화수소(H2O2)를 생성한다. 과산화수소는 카탈레이스(Catalase)에 의해 즉시 분해되며 열을 방출한다.

둘째로, 퍼옥시좀의 β 산화는 긴사슬 지방산, 혹은 가지 사슬 지방산에 큰 활성을 가진다. 이러한 지방산은 유제품, 생선 등의 식이를 통해 유입된다. 이 과정은 은 주로 간 퍼옥시좀에서 일어난다. 퍼옥시좀에서 사슬 길이가 짧아진 지방산은 퍼옥시좀 밖으로 나간 후 다시 미토콘드리아로 유입된다.
5. ω 산화
소포체(Endoplasmic reticulum, 세포질세망이라고도 함)에서는 ω 산화가 일어난다. ω 산화는 일반적인 지방산 분해 경로는 아니지만, 돌연변이 혹은 카니틴 결핍에 의해 β 산화가 수행될 수 없을 때 매우 중요해진다.
ω 산화의 첫 단계는 ω 탄소의 메틸기(-CH3)를 카복실기로 바꾸는 것이다. 이 과정은 세 번의 산화(메틸→알코올→알데하이드→카복실산)를 거쳐 일어난다.

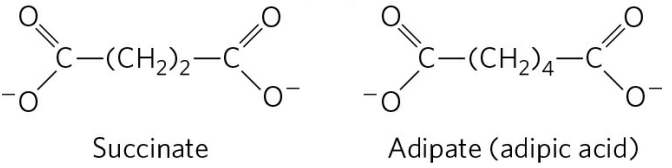
양쪽 끝이 모두 카복실기인 화합물은 카니틴 없이도 미토콘드리아 내부로 유입될 수 있고, 미토콘드리아 내부에서 양 쪽이 동시에 β 산화가 진행된다. 결과적으로 4탄소 화합물인 석신산, 6탄소 화합물인 아디프산 등이 생성된다.
6. α 산화
가지 사슬 지방산, 특히 유제품 등에 함유된 파이탄산(Phytanic acid)은 β 탄소에 메틸기가 결합해 있어 β 산화가 불가능하다. 따라서 파이탄산은 퍼옥시좀에 들어가 α 산화에 의해 분해된다. α 산화는 파이탄산의 메틸기가 β 탄소에 존재하지 않도록 α 탄소를 CO2의 형태로 제거하고, 이후 β 산화를 진행해 메틸기가 결합된 아세틸-CoA, 프로피오닐-CoA를 산물로 내보낸다. 자세한 반응 기전은 다음 그림을 살펴보자.

지방산의 분해는 매우 복잡한 과정이다. 이 부분을 공부하는 데 있어 지방산의 이화 과정을 직접 그려보면 큰 도움이 될 것이다.

'생화학 > 생화학 (하)' 카테고리의 다른 글
| [생화학] 아미노기 대사 (Metabolism of Amino Groups) (0) | 2022.08.21 |
|---|---|
| [생화학] 케톤체 (Ketone Bodies) (0) | 2022.08.17 |
| [생화학] 지방의 소화, 동원 및 운반 (Digestion, Mobilization, and Transport of Fats) (0) | 2022.08.10 |
| [생화학] 시트르산 회로 (The Citric Acid Cycle) (0) | 2022.08.09 |
| [생화학] 아세틸-CoA의 생성 (Production of Acetyl-CoA) (0) | 2022.08.06 |