지방[Fat, 중성 지방(Neutral fat) 혹은 트라이아실글리세롤(Triacylglycerol)이라 불리기도 함]은 지질(Lipid)의 한 종류다. 지질이란 물에 녹지 않는 불용성 생체 분자를 통칭한다. 그 중에서도 지방은 세포 내 에너지의 저장 역할을 담당한다.
지방은 한 분자의 글리세롤(Glycerol)과 세 분자의 지방산(Fatty acid)이 결합해 이루어진다. 글리세롤은 세 개의 수산화기를 갖는 3탄소 화합물이다.
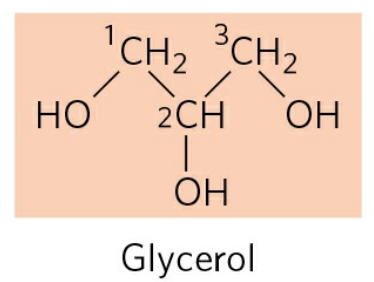
지방산은 카복실기를 가진 긴 사슬 탄화수소를 일컫는다. 생체 지방산의 탄소 수는 대개 짝수로 증가한다.

탄화수소 사슬이 모두 단일 결합으로 이루어진 경우 이를 탄소가 수소로 포화되었다는 뜻에서 포화지방산(Saturated fatty acid)이라 한다. 반면, 하나 이상의 이중 결합을 포함할 경우 이를 불포화지방산(Unsaturated fatty acid)이라 한다. 불포화지방산은 다시 이중 결합의 방향에 따라 트랜스 지방산(Trans fatty acid)과 시스 지방산(Cis fatty acid)으로 분류할 수 있다. 생물 유래 지방산은 대부분 시스 지방산이다. 둘의 차이점은 지방산의 산화에서 보다 자세히 다뤄보도록 하자.
지방산의 탄소 번호는 카복실기의 탄소가 1번임을 기준으로 한다. 위 그림의 지방산은 탄소가 18개다. 여기서 이중결합의 위치는 9번과 10번 탄소 사이에 있는데, 이를 간단히 18:1(Δ9)라 쓴다. 18은 탄소의 개수, 1은 이중결합의 개수이다.
지방산과 글리세롤의 결합은 카복실기와 수산화기의 결합이다. 이때 한 분자의 물이 빠져나가고, 이러한 결합 방식을 에스터(Ester) 결합이라 한다. 글리세롤에 결합한 세 지방산이 모두 같을 경우 단순 트라이아실글리세롤, 종류가 다를 경우 혼합 트라이아실글리세롤이라 한다. 생체 내 대부분의 트라이아실글리세롤은 혼합형이다.

트라이아실글리세롤에서 지방산은 일종의 아실기(R-C=O-)가 된다. 트라이아실글리세롤의 명칭은 세 개의 아실기가 결합한 데서 유래한다.
지방은 생체 내에서 어떻게 대사되는가? 이번 글에서는 식이, 저장 지방이 어떻게 대사 경로로 들어가는지에 대해 알아보도록 하자.

1. 지방 대사 개요
지방은(특히 지방산)은 많은 에너지를 갖는 고에너지 화합물이다. 많은 장기와 조직에서 지방산의 산화는 중심이 되는 에너지 생산 경로다. 지방산과 유리된 글리세롤은 쉽게 당분해 경로로 들어갈 수 있다. 지방산은 크게 4가지 경로에서 공급된다;
① 식이 지방 : 식이성 트라이아실글리세롤은 소화 경로를 거친다.
② 저장 지방 : 지방세포의 지질 방울로 저장된 지방은 호르몬에 의해 동원된다.
③ 합성 지방 : 간에서 과잉 섭취된 포도당은 지방으로 전환될 수 있다.
④ 분해 지방 : 자가포식(Autophagy) 작용을 통해 지방을 얻을 수 있다.
다양한 경로에서 유래된 지방산은 이화되며 다량의 아세틸-CoA를 생성하고, 아세틸-CoA는 시트르산 회로로 들어가 ATP를 합성한다. 지방은 고도로 환원된 화합물이기 때문에 많은 에너지를 함유할 수 있는데, 동시에 고도로 환원되었기 때문에 지방은 물에 녹을 수 없다. 불용성 화합물은 혈액을 통해 수송될 수 없으므로 지방은 특별한 운반 경로를 필요로 한다.
2. 지방의 소화
식이성 지방은 쓸개즙(Bile salt)과 지질분해효소(Lipase, 라이페이스)에 의해 소장에서 소화된다. 우선 쓸개즙이 식이성 지방 분자를 유화시켜 마이셀(Micelle)을 형성하면, 이들은 라이페이스에 의해 에스터 결합이 끊겨 모노아실글리세롤, 다이아실글리세롤, 유리 지방산, 글리세롤 등으로 분리된다.

라이페이스에 의한 분해산물은 소장 점막의 상피세포로 확산되고, 이후 다시 트라이아실글리세롤로 합쳐진다. 합쳐진 트라이아실글리세롤은 콜레스테롤(Cholesterol), 아포지단백질(Apolipoprotein)과 함께 암죽미립(Chylomicron)이라 불리는 거대한 구(Sphere)를 형성한다.
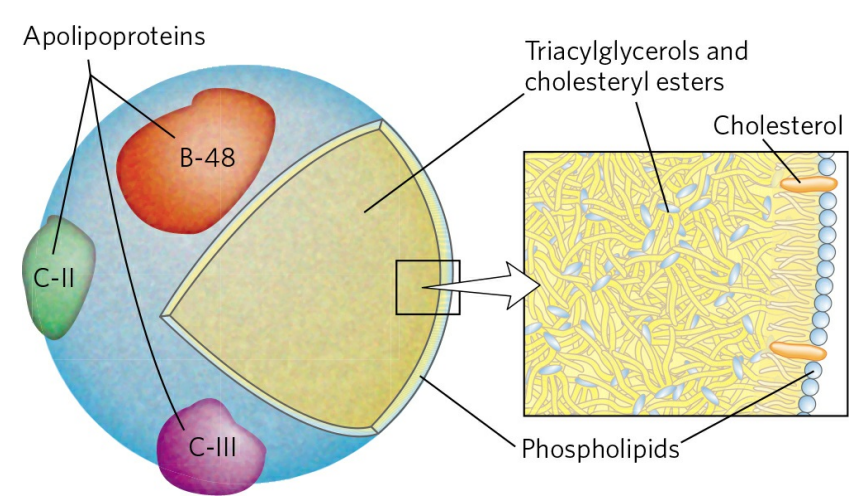
암죽미립의 내부는 트라이아실글리세롤로 꽉 차있고, 이를 인지질과 콜레스테롤로 된 막이 감싼다. 막에는 아포지단백질이 결합해있다. 아포지단백질은 일종의 신호 역할을 한다. 예를 들어, C-Ⅱ 아포지단백질을 가진 암죽미립은 창자 점막에서 림프계(Lymphatic system)로 이동해 걸러진 후, 혈중으로 돌아가 근육과 지방조직으로 운반된다. 운반받는 조직의 세포 밖에는 지단백질 지질분해효소(Lipoprotein lipase)가 존재하는데, 이 효소는 C-Ⅱ를 인식해 활성화되어 암죽미립을 분해하고 트라이아실글리세롤의 에스터결합을 모두 끊어 지방산과 글리세롤로 유리한다.
지방산과 글리세롤은 조직 세포 내로 운반된다. 근육에서는 지방산이 산화되어 에너지로 사용되고, 지방조직(Adipocyte)에서는 유입된 지방산을 재에스터화해 트라이아실글리세롤로 저장한다.

트라이아실글리세롤이 지단백질 지질분해효소에 의해 분해되고 남은 암죽미립 잔해(아포지단백질, 인지질, 콜레스테롤 등)는 혈액을 따라 간으로 운반된다.
3. 지방의 동원
지방은 지방세포에 지질 방울(Lipid droplet)의 형태로 저장되어 있다. 지질 방울은 암죽미립과 유사하게 단층의 인지질 막 내부에 다량의 트라이아실글리세롤이 저장되어 있고, 인지질 표면은 페리리핀(Perilipins)이라 불리는 단백질로 싸여있다. 페리리핀은 지질방울이 아무때나 소모되지 않도록 방울을 보호하는 역할을 한다. 저에너지 상태에서 특정한 호르몬이 지방세포에 에너지 공급 신호를 보내면, 지방조직의 트라이아실글리세롤은 지질 방울로부터 동원된다.
저에너지 상태를 알리는 호르몬은 글루카곤과 에피네프린이다. 이들은 지방세포의 아데닐산 고리화효소(Adenylyl cyclase)를 활성화해 cAMP를 생산하고, cAMP 의존 단백질 인산화효소 A(PKA)가 여러 지방분해효소들을 촉진한다. 이는 글루카곤이 글리코겐의 분해를 촉진하는 연쇄증폭 반응과 매우 유사하다.
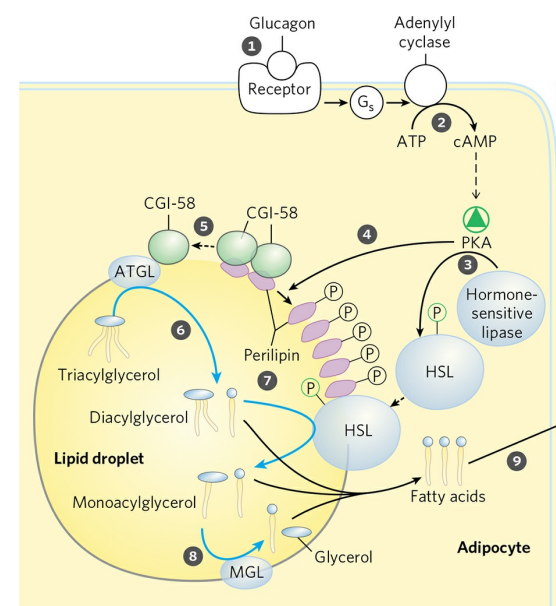
PKA는 지방분해효소뿐만 아니라 페리리핀 분자도 인산화한다. 인산화된 페리리핀은 라이페이스가 지질 방울 내부의 트라이아실글리세롤에 접근할 수 있도록 돕는다.
유리 지방산은 혈액을 통해 수송되어야 하지만, 지방산은 불용성이다. 때문에 지방산은 혈청 알부민(Serum albumin)이라 불리는 단백질에 결합해 수송된다. 알부민 한 분자 당 10개까지의 지방산이 결합할 수 있다. 표적 조직에 도착하면 알부민에서 지방산이 분리되고, 지방산은 세포 내로 이동하여 에너지원으로 이용된다.
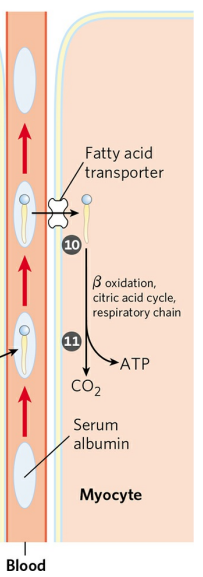
유리 글리세롤은 글리세롤 인산화효소(Glycerol kinase)에 의해 3-인산 글리세롤로 전환된다. 3-인산 글리세롤은 당분해 경로로 들어간다.
4. 지방산의 운반
지방산의 산화는 미토콘드리아 내부에서 일어난다. 탄소 12개 이하의 지방산은 별다른 수송체 없이 미토콘드리아 내부로 유입될 수 있지만, 일반적으로 대부분의 지방산을 차지하는 탄소 14개 이상의 지방산은 미토콘드리아막을 통과할 수 없다. 때문에 이들은 카니틴 왕복통로(Carnitine shuttle)라 불리는 특별한 운반 기전을 이용한다.
카니틴 왕복통로의 첫반응은 아실-CoA 합성효소(Acyl-CoA synthetase)에 의한 지방산과 CoA의 합성이다. 이 반응은 1분자의 ATP를 소모한다. 지방산은 일종의 아실기라 했던 것을 기억하자.
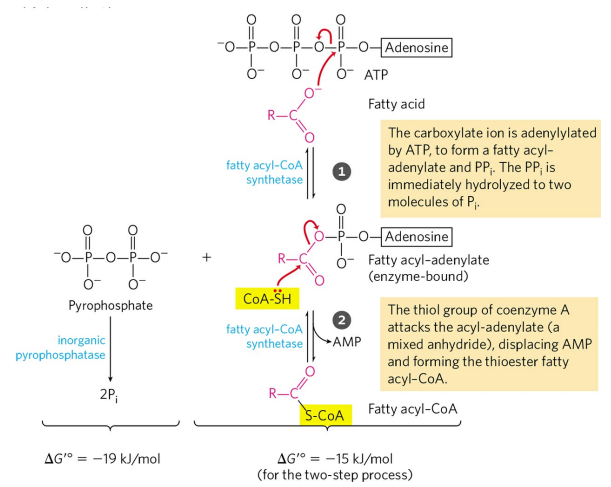
그 후, 카니틴 아실기전달효소 1(Carnitine acyltransferase 1, CPT1)에 의해 CoA가 카니틴으로 치환되어 아실-카니틴 중합체를 형성한다. 아실-카니틴 중합체가 미토콘드리아 외부에서 만들어지는지 막 사이 공간에서 만들어지는지는 아직 밝혀지지 않았다.
이후, 아실-카니틴 중합체는 아실-카니틴/카니틴 공수송체(Acyl-carnitine/carnitine cotransporter)에 의해 미토콘드리아 내부로 유입된다. 동시에 아무것도 결합하지 않은 카니틴이 미토콘드리아 외부로 방출된다.
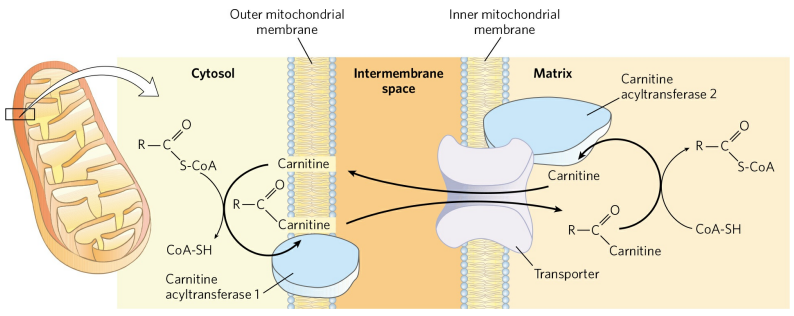
미토콘드리아 내부로 유입된 아실-카니틴은 카니틴 아실기전달효소 2(CPT2)에 의해 다시 아실-CoA로 전환된다. 이러한 복잡한 수송은 두 영역(미토콘드리아 내부와 세포질)에서 아실-CoA의 운명을 다르게 하기 위함이다. 미토콘드리아 내부에서 아실-CoA는 에너지의 생산을 위해 산화되고, 세포질에서 아실-CoA는 지질의 생합성을 위해 이용된다.
지금까지 식이성 지방, 저장 지방이 어떻게 미토콘드리아 내부로 유입되는지에 대해 알아보았다. 미토콘드리아 내부로 유입되는 카니틴 통로 반응은 지방산 산화의 속도 제한 단계다. 일단 아실-CoA가 미토콘드리아 내부로 유입되면, 지방산의 산화는 매우 빠르게 일어난다. 지방산은 어떻게 산화되는가? 이에 대해서는 다음 글에서 알아보도록 하자.
미토콘드리아 내부로 유입된 지방산이 어떻게 산화되는지에 대해서는 다음 글에서 알아보도록 하자.

'생화학 > 생화학 (하)' 카테고리의 다른 글
| [생화학] 케톤체 (Ketone Bodies) (0) | 2022.08.17 |
|---|---|
| [생화학] 지방산의 산화 (Oxidation of Fatty Acids) (3) | 2022.08.13 |
| [생화학] 시트르산 회로 (The Citric Acid Cycle) (0) | 2022.08.09 |
| [생화학] 아세틸-CoA의 생성 (Production of Acetyl-CoA) (0) | 2022.08.06 |
| [생화학] 글리코겐 대사의 조절 (Regulation of The Metabolism of Glycogen) (0) | 2022.08.03 |



